

Reverse engineering synthetic antiviral amyloids
- PMID: 32504029
- PMCID: PMC7275043
- DOI: 10.1038/s41467-020-16721-8
Reverse engineering synthetic antiviral amyloids
Erratum in
-
Author Correction: Reverse engineering synthetic antiviral amyloids.Nat Commun. 2023 Jun 13;14(1):3492. doi: 10.1038/s41467-023-39293-9. Nat Commun. 2023. PMID: 37311856 Free PMC article. No abstract available.
Abstract
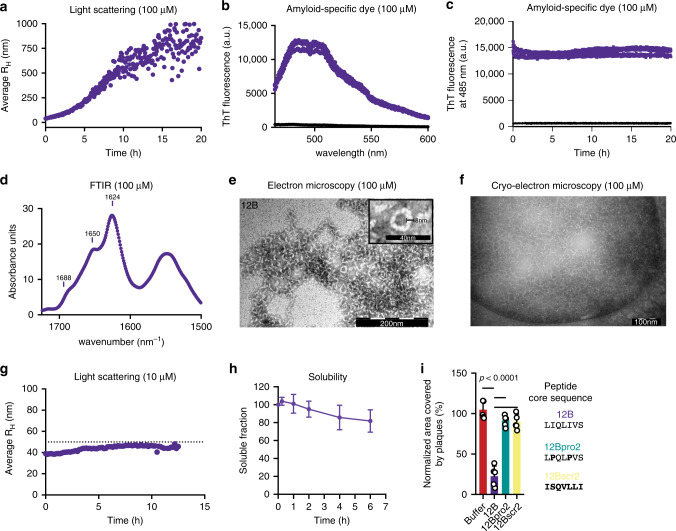
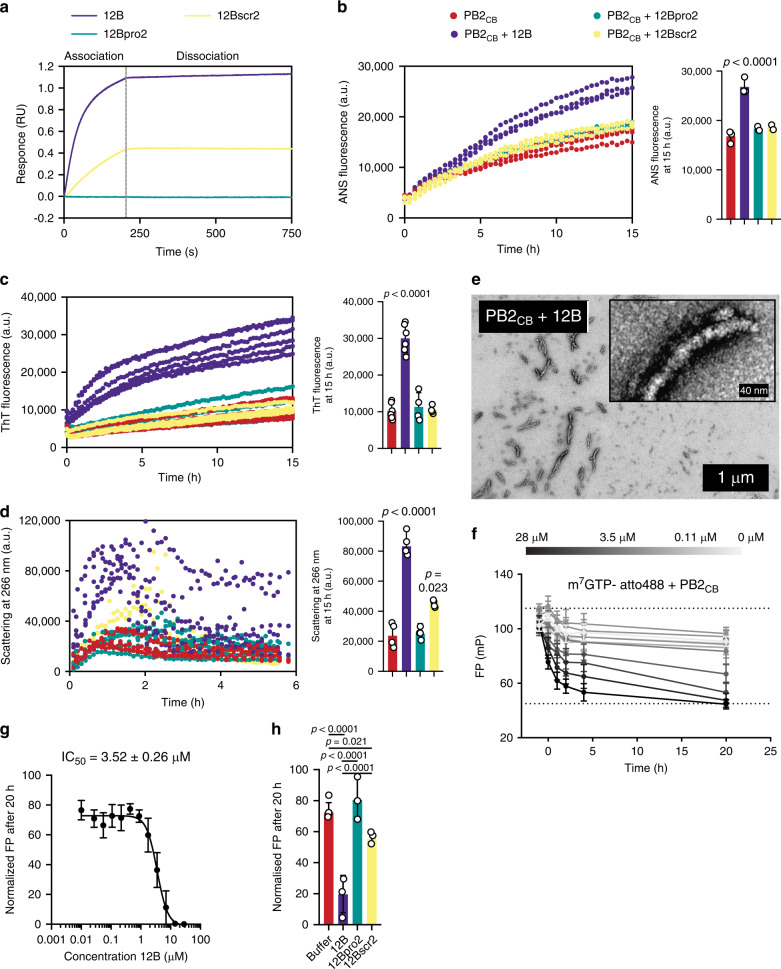
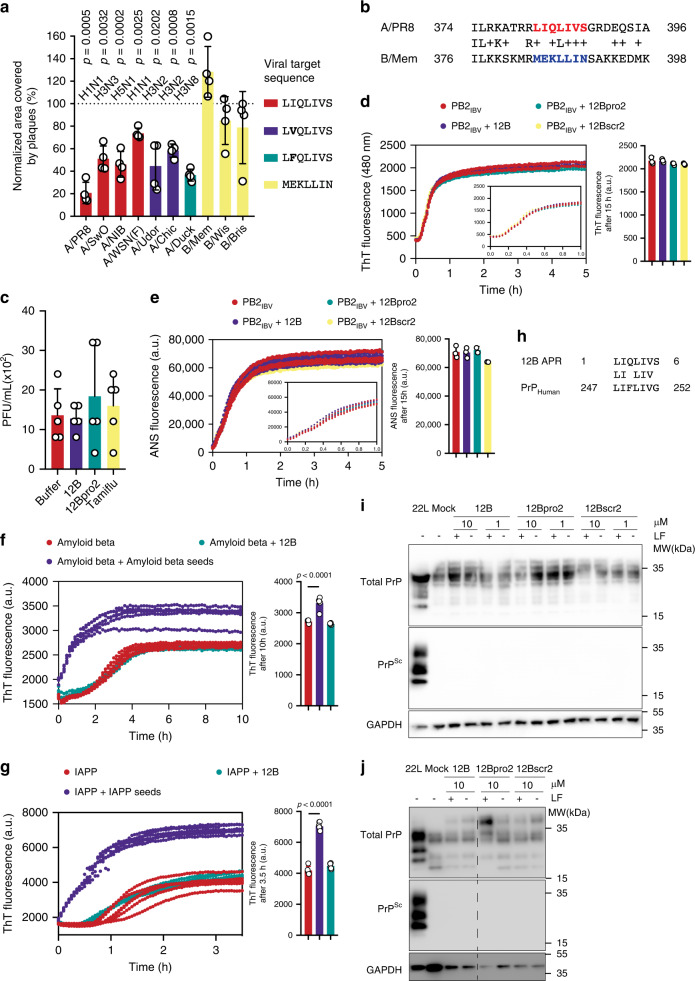
Human amyloids have been shown to interact with viruses and interfere with viral replication. Based on this observation, we employed a synthetic biology approach in which we engineered virus-specific amyloids against influenza A and Zika proteins. Each amyloid shares a homologous aggregation-prone fragment with a specific viral target protein. For influenza we demonstrate that a designer amyloid against PB2 accumulates in influenza A-infected tissue in vivo. Moreover, this amyloid acts specifically against influenza A and its common PB2 polymorphisms, but not influenza B, which lacks the homologous fragment. Our model amyloid demonstrates that the sequence specificity of amyloid interactions has the capacity to tune amyloid-virus interactions while allowing for the flexibility to maintain activity on evolutionary diverging variants.
Conflict of interest statement
The authors declare the following competing interests: J.S. and F.R. are listed as inventors of patents held by their hold institution VIB, covering the peptides described in this manuscript, and which are licensed to Aelin Therapeutics (Leuven, Belgium), of which J.S. and F.R. are the scientific founders. All other authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
