

Biallelic variants in the RNA exosome gene EXOSC5 are associated with developmental delays, short stature, cerebellar hypoplasia and motor weakness
- PMID: 32504085
- PMCID: PMC7399534
- DOI: 10.1093/hmg/ddaa108
Biallelic variants in the RNA exosome gene EXOSC5 are associated with developmental delays, short stature, cerebellar hypoplasia and motor weakness
Abstract
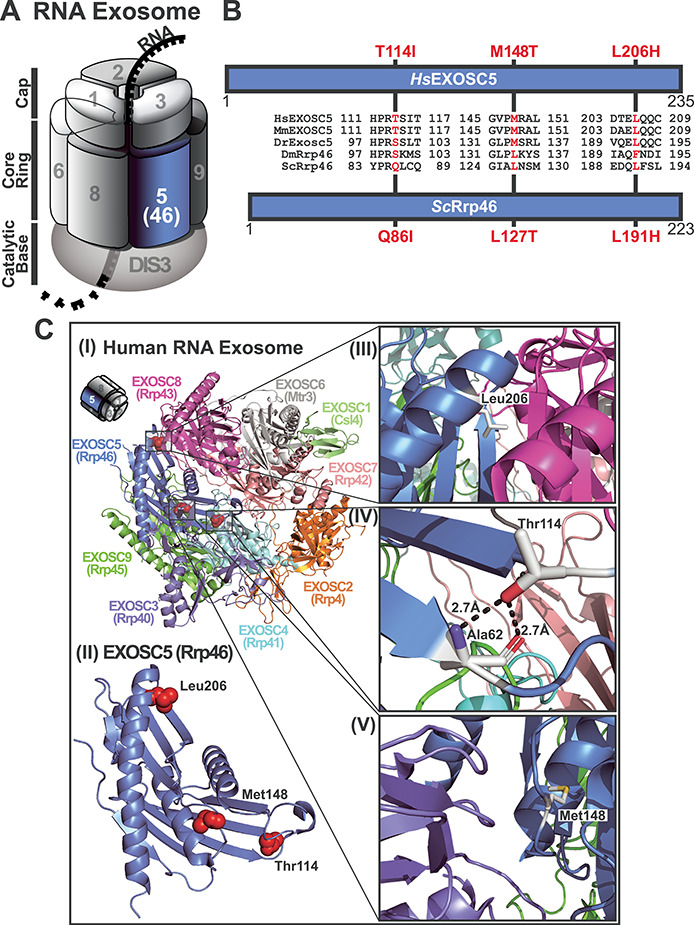
The RNA exosome is an essential ribonuclease complex required for processing and/or degradation of both coding and non-coding RNAs. We identified five patients with biallelic variants in EXOSC5, which encodes a structural subunit of the RNA exosome. The clinical features of these patients include failure to thrive, short stature, feeding difficulties, developmental delays that affect motor skills, hypotonia and esotropia. Brain MRI revealed cerebellar hypoplasia and ventriculomegaly. While we ascertained five patients, three patients with distinct variants of EXOSC5 were studied in detail. The first patient had a deletion involving exons 5-6 of EXOSC5 and a missense variant, p.Thr114Ile, that were inherited in trans, the second patient was homozygous for p.Leu206His and the third patient had paternal isodisomy for chromosome 19 and was homozygous for p.Met148Thr. The additional two patients ascertained are siblings who had an early frameshift mutation in EXOSC5 and the p.Thr114Ile missense variant that were inherited in trans. We employed three complementary approaches to explore the requirement for EXOSC5 in brain development and assess consequences of pathogenic EXOSC5 variants. Loss of function for exosc5 in zebrafish results in shortened and curved tails/bodies, reduced eye/head size and edema. We modeled pathogenic EXOSC5 variants in both budding yeast and mammalian cells. Some of these variants cause defects in RNA exosome function as well as altered interactions with other RNA exosome subunits. These findings expand the number of genes encoding RNA exosome subunits linked to human disease while also suggesting that disease mechanism varies depending on the specific pathogenic variant.
© The Author(s) 2020. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures






References
-
- Brouwer R., Allmang C., Raijmakers R., van Aarssen Y., Egberts W.V., Petfalski E., van Venrooij W.J., Tollervey D. and Pruijn G.J. (2001) Three novel components of the human exosome. J. Biol. Chem., 276, 6177–6184. - PubMed
-
- Mitchell P., Petfalski E. and Tollervey D. (1996) The 3′ end of yeast 5.8S rRNA is generated by an exonuclease processing mechanism. Genes Dev., 10, 502–513. - PubMed
-
- Mitchell P., Petfalski E., Shevchenko A., Mann M. and Tollervey D. (1997) The exosome: a conserved eukaryotic RNA processing complex containing multiple 3′→5′ exoribonucleases. Cell, 91, 457–466. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
