

Disrupted inhibitory plasticity and homeostasis in Fragile X syndrome
- PMID: 32512151
- PMCID: PMC7959200
- DOI: 10.1016/j.nbd.2020.104959
Disrupted inhibitory plasticity and homeostasis in Fragile X syndrome
Abstract
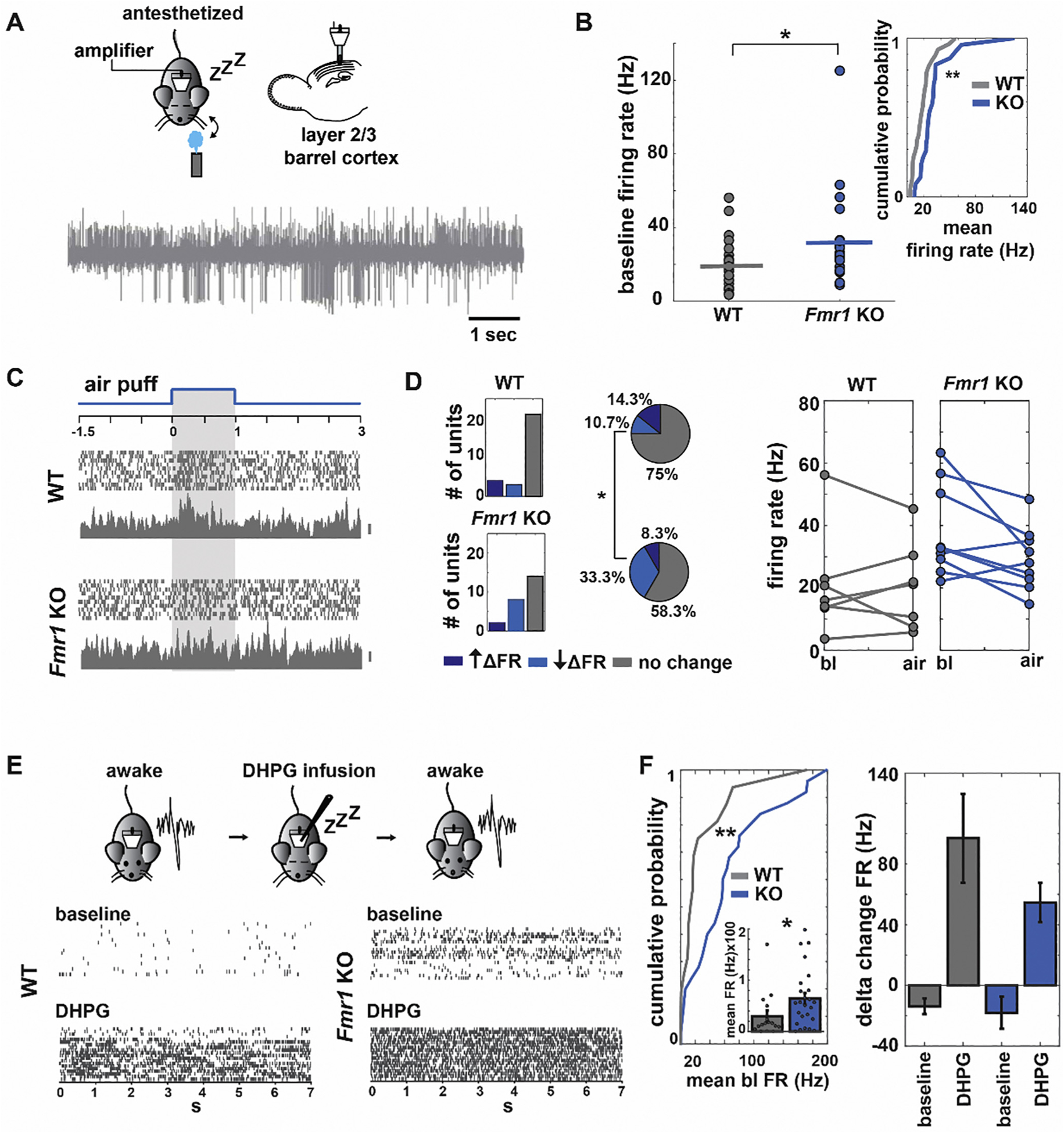
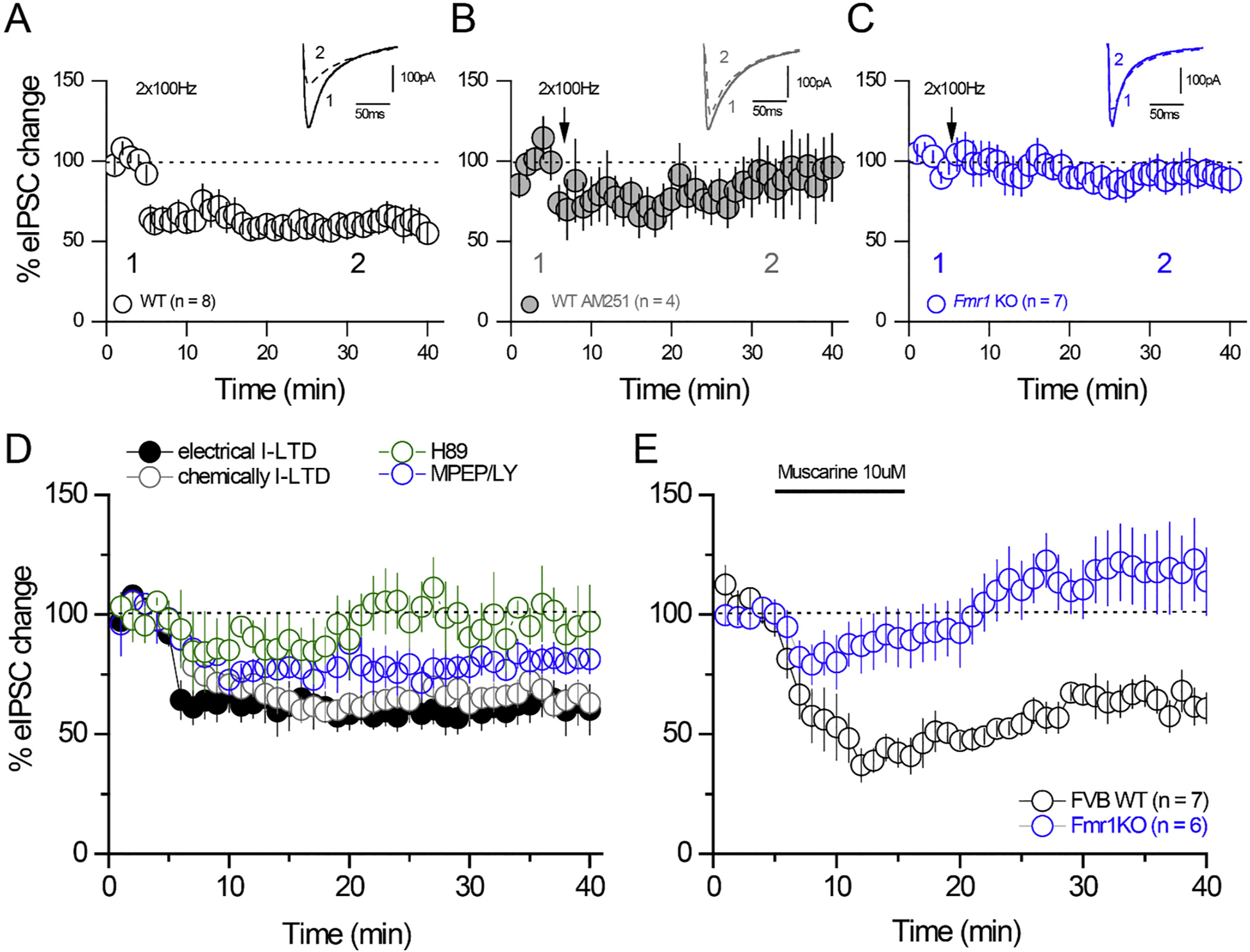
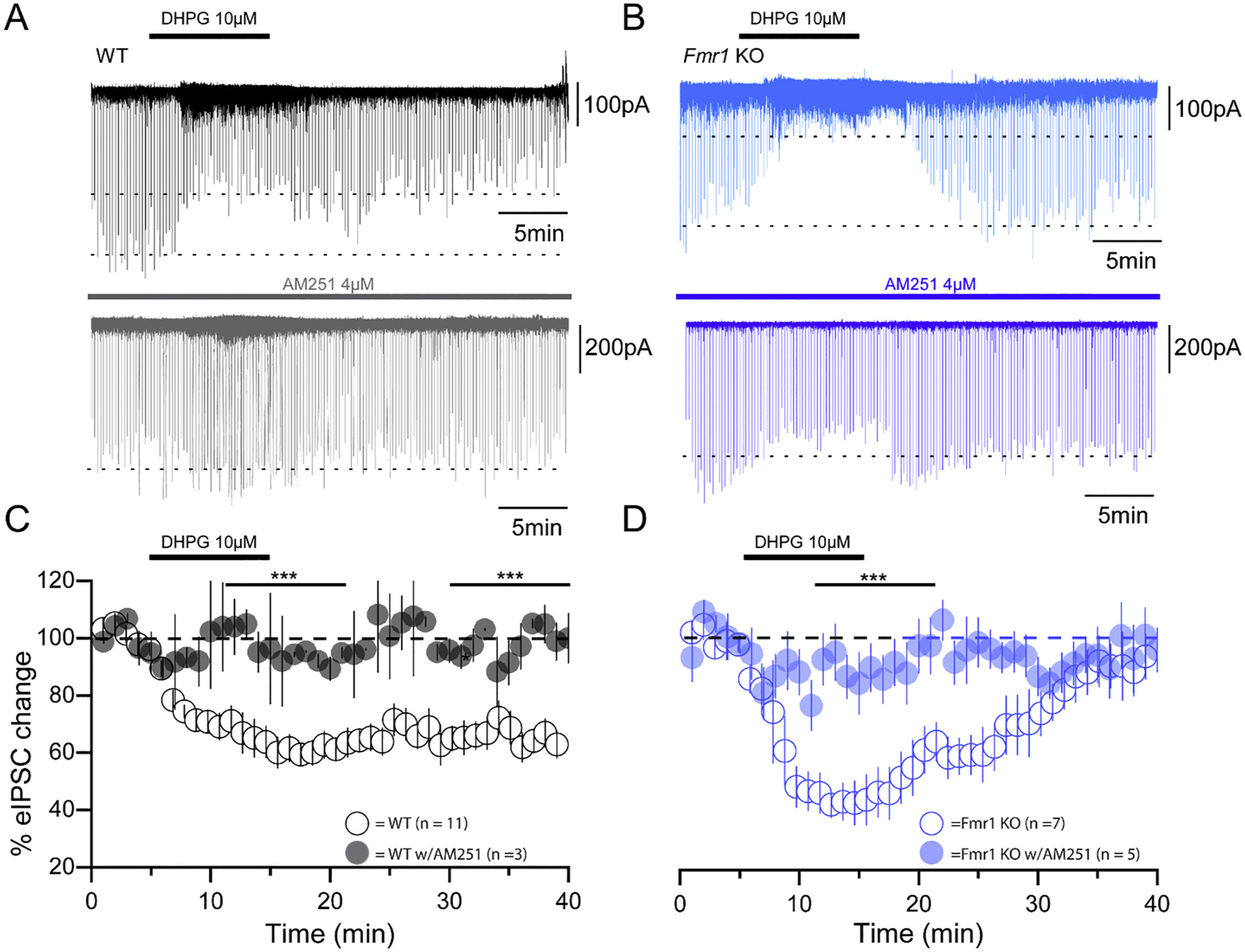
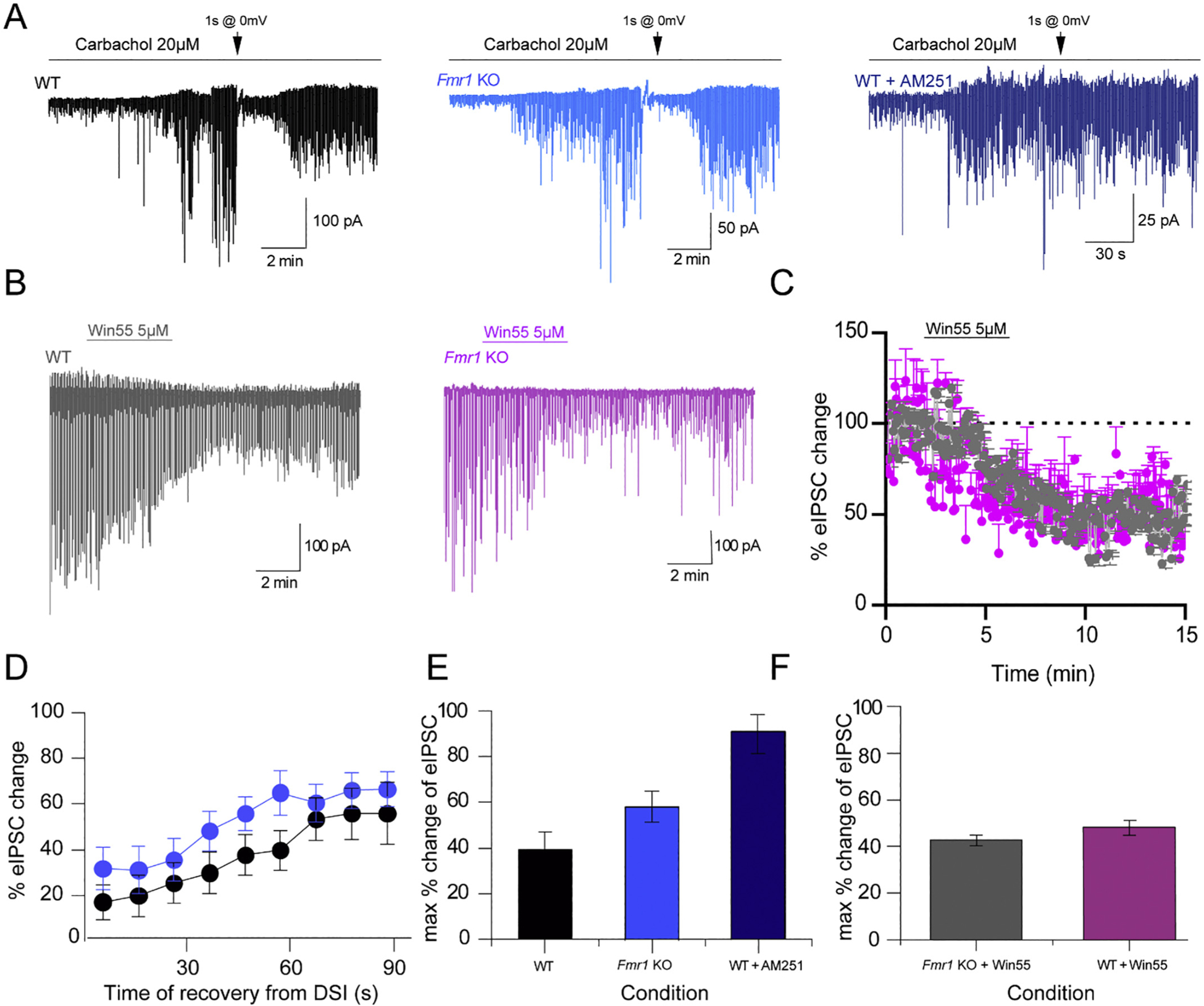
Fragile X Syndrome (FXS) is a neurodevelopmental disorder instigated by the absence of a key translation regulating protein, Fragile X Mental Retardation Protein (FMRP). The loss of FMRP in the CNS leads to abnormal synaptic development, disruption of critical periods of plasticity, and an overall deficiency in proper sensory circuit coding leading to hyperexcitable sensory networks. However, little is known about how this hyperexcitable environment affects inhibitory synaptic plasticity. Here, we show that in vivo layer 2/3 of the primary somatosensory cortex of the Fmr1 KO mouse exhibits basal hyperexcitability and an increase in neuronal firing rate suppression during whisker activation. This aligns with our in vitro data that indicate an increase in GABAergic spontaneous activity, a faulty mGluR-mediated inhibitory input and impaired inhibitory plasticity processes. Specifically, we find that mGluR activation sensitivity is overall diminished in the Fmr1 KO mouse leading to both a decreased spontaneous inhibitory postsynaptic input to principal cells and a disrupted form of inhibitory long-term depression (I-LTD). These data suggest an adaptive mechanism that acts to homeostatically counterbalance the cortical hyperexcitability observed in FXS.
Keywords: Cortex; Fragile X syndrome; Inhibitory neurotransmission; Interneurons; Plasticity.
Copyright © 2020. Published by Elsevier Inc.
Figures

Similar articles
-
Impaired inhibitory control of cortical synchronization in fragile X syndrome.J Neurophysiol. 2011 Nov;106(5):2264-72. doi: 10.1152/jn.00421.2011. Epub 2011 Jul 27. J Neurophysiol. 2011. PMID: 21795626 Free PMC article.
-
Impairment in the homeostatic recruitment of layer 5/6 neurons following whisker stimulation in Fmr1 KO mice.Neurobiol Dis. 2025 Apr;207:106837. doi: 10.1016/j.nbd.2025.106837. Epub 2025 Feb 10. Neurobiol Dis. 2025. PMID: 39938578 Free PMC article.
-
Abnormal presynaptic short-term plasticity and information processing in a mouse model of fragile X syndrome.J Neurosci. 2011 Jul 27;31(30):10971-82. doi: 10.1523/JNEUROSCI.2021-11.2011. J Neurosci. 2011. PMID: 21795546 Free PMC article.
-
Fragile X syndrome: the GABAergic system and circuit dysfunction.Dev Neurosci. 2011;33(5):349-64. doi: 10.1159/000329420. Epub 2011 Sep 21. Dev Neurosci. 2011. PMID: 21934270 Free PMC article. Review.
-
BDNF in fragile X syndrome.Neuropharmacology. 2014 Jan;76 Pt C:729-36. doi: 10.1016/j.neuropharm.2013.05.018. Epub 2013 May 29. Neuropharmacology. 2014. PMID: 23727436 Review.
Cited by
-
Glucocorticoid regulation and neuroanatomy in fragile x syndrome.J Psychiatr Res. 2021 Feb;134:81-88. doi: 10.1016/j.jpsychires.2020.12.015. Epub 2020 Dec 11. J Psychiatr Res. 2021. PMID: 33373777 Free PMC article.
-
Neurodevelopment and early pharmacological interventions in Fragile X Syndrome.Front Neurosci. 2023 Aug 2;17:1213410. doi: 10.3389/fnins.2023.1213410. eCollection 2023. Front Neurosci. 2023. PMID: 37599992 Free PMC article. Review.
-
Compensatory Regulation of Excitation/Inhibition Balance in the Ventral Hippocampus: Insights from Fragile X Syndrome.Biology (Basel). 2025 Mar 31;14(4):363. doi: 10.3390/biology14040363. Biology (Basel). 2025. PMID: 40282228 Free PMC article. Review.
-
Increased Pain Symptomatology Among Females vs. Males With Fragile X-Associated Tremor/Ataxia Syndrome.Front Psychiatry. 2022 Jan 20;12:762915. doi: 10.3389/fpsyt.2021.762915. eCollection 2021. Front Psychiatry. 2022. PMID: 35126193 Free PMC article.
-
Increased Inhibition May Contribute to Maintaining Normal Network Function in the Ventral Hippocampus of a Fmr1-Targeted Transgenic Rat Model of Fragile X Syndrome.Brain Sci. 2023 Nov 17;13(11):1598. doi: 10.3390/brainsci13111598. Brain Sci. 2023. PMID: 38002556 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
