

Differential Coassembly of α1-GABAARs Associated with Epileptic Encephalopathy
- PMID: 32513829
- PMCID: PMC7363476
- DOI: 10.1523/JNEUROSCI.2748-19.2020
Differential Coassembly of α1-GABAARs Associated with Epileptic Encephalopathy
Abstract
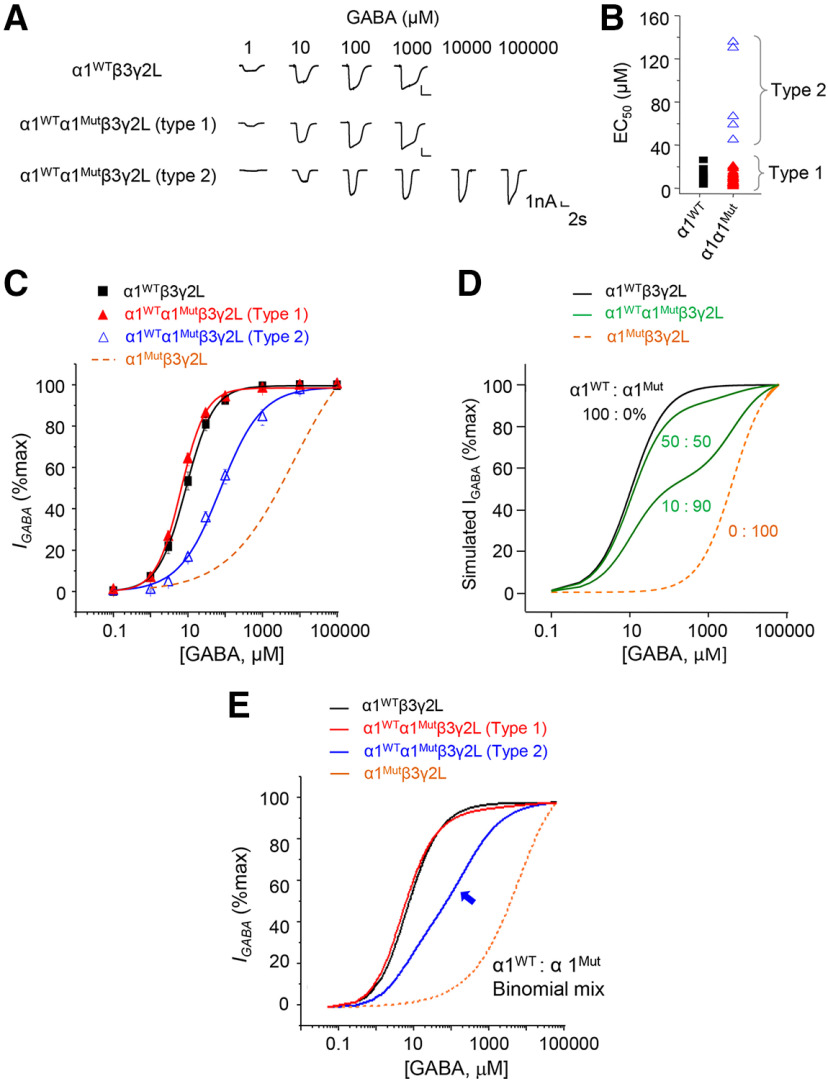
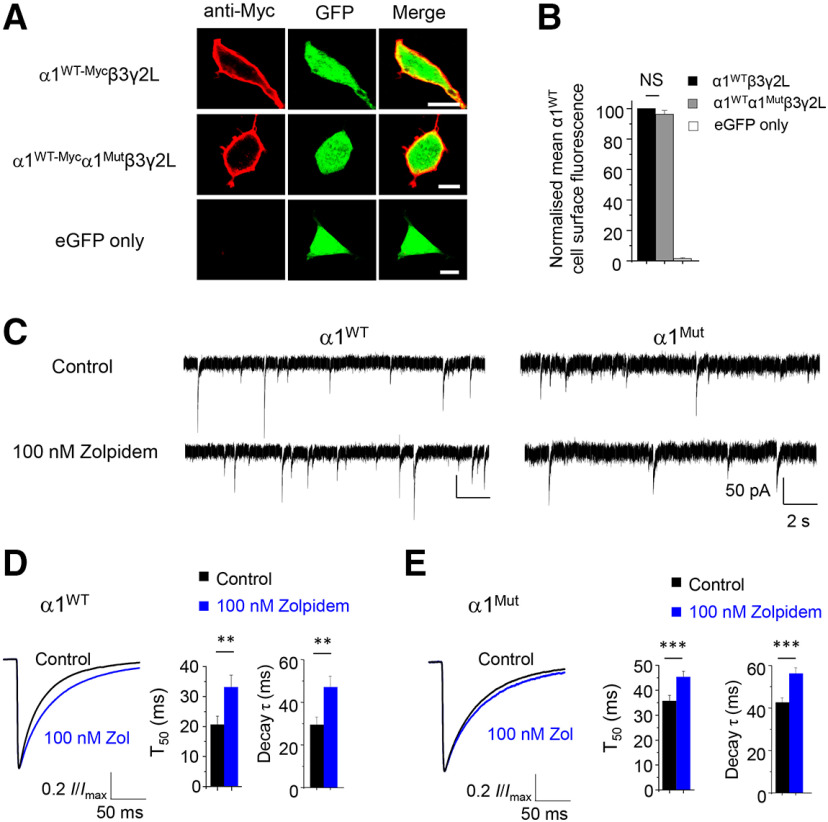
GABAA receptors (GABAARs) are profoundly important for controlling neuronal excitability. Spontaneous and familial mutations to these receptors feature prominently in excitability disorders and neurodevelopmental deficits following disruption to GABA-mediated inhibition. Recent genotyping of an individual with severe epilepsy and Williams-Beuren syndrome identified a frameshifting de novo variant in a major GABAAR gene, GABRA1 This truncated the α1 subunit between the third and fourth transmembrane domains and introduced 24 new residues forming the mature protein, α1Lys374Serfs*25 Cell surface expression of mutant murine GABAARs is severely impaired compared with WT, due to retention in the endoplasmic reticulum. Mutant receptors were differentially coexpressed with β3, but not with β2, subunits in mammalian cells. Reduced surface expression was reflected by smaller IPSCs, which may underlie the induction of seizures. The mutant does not have a dominant-negative effect on native neuronal GABAAR expression since GABA current density was unaffected in hippocampal neurons, although mutant receptors exhibited limited GABA sensitivity. To date, the underlying mechanism is unique for epileptogenic variants and involves differential β subunit expression of GABAAR populations, which profoundly affected receptor function and synaptic inhibition.SIGNIFICANCE STATEMENT GABAARs are critical for controlling neural network excitability. They are ubiquitously distributed throughout the brain, and their dysfunction underlies many neurologic disorders, especially epilepsy. Here we report the characterization of an α1-GABAAR variant that results in severe epilepsy. The underlying mechanism is structurally unusual, with the loss of part of the α1 subunit transmembrane domain and part-replacement with nonsense residues. This led to compromised and differential α1 subunit cell surface expression with β subunits resulting in severely reduced synaptic inhibition. Our study reveals that disease-inducing variants can affect GABAAR structure, and consequently subunit assembly and cell surface expression, critically impacting on the efficacy of synaptic inhibition, a property that will orchestrate the extent and duration of neuronal excitability.
Keywords: GABA-A receptor; epilepsy; inhibition; synaptic transmission; α subunit variant.
Copyright © 2020 the authors.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical