

Comparative tRNA sequencing and RNA mass spectrometry for surveying tRNA modifications
- PMID: 32514182
- PMCID: PMC8172280
- DOI: 10.1038/s41589-020-0558-1
Comparative tRNA sequencing and RNA mass spectrometry for surveying tRNA modifications
Abstract
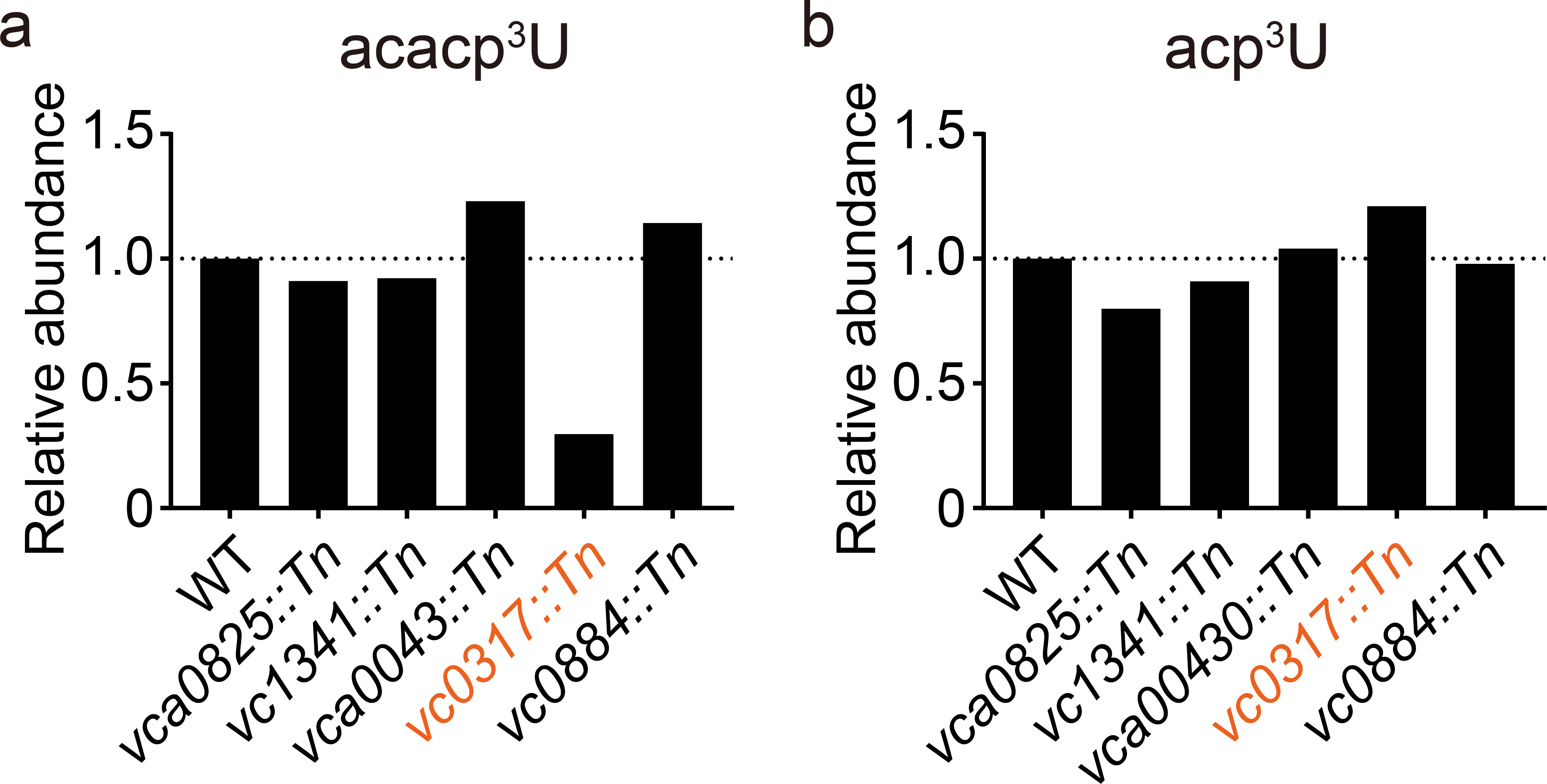
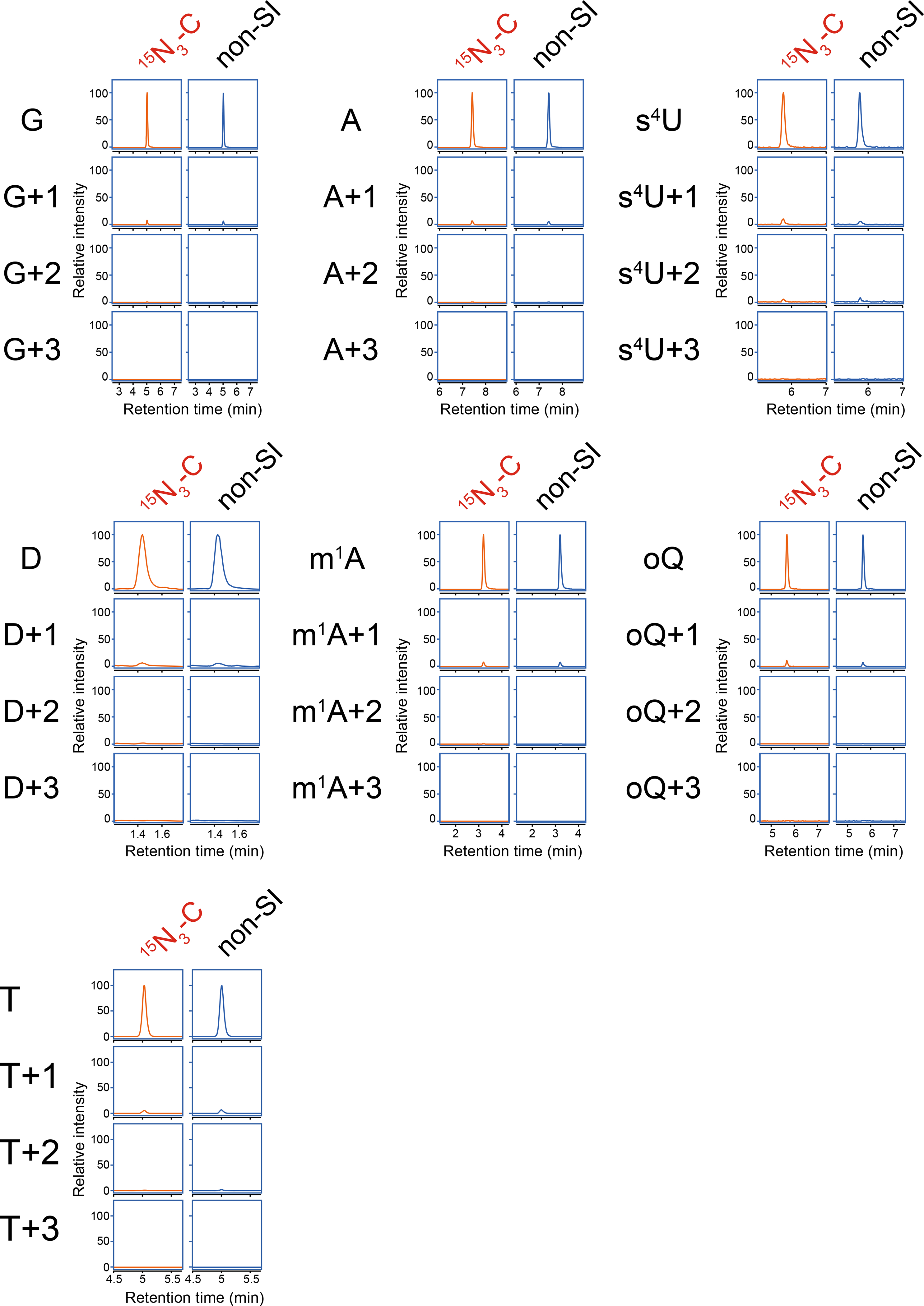
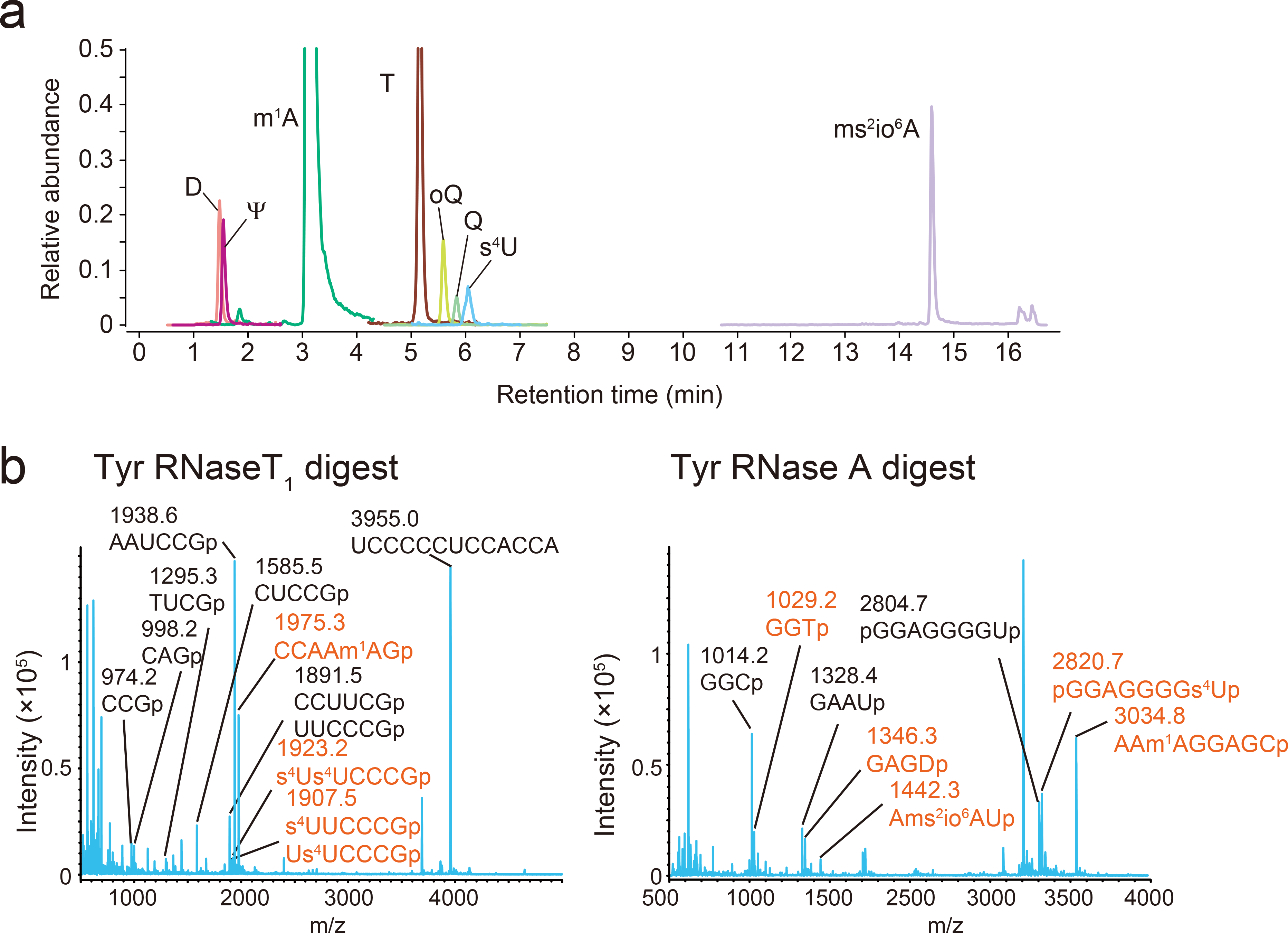
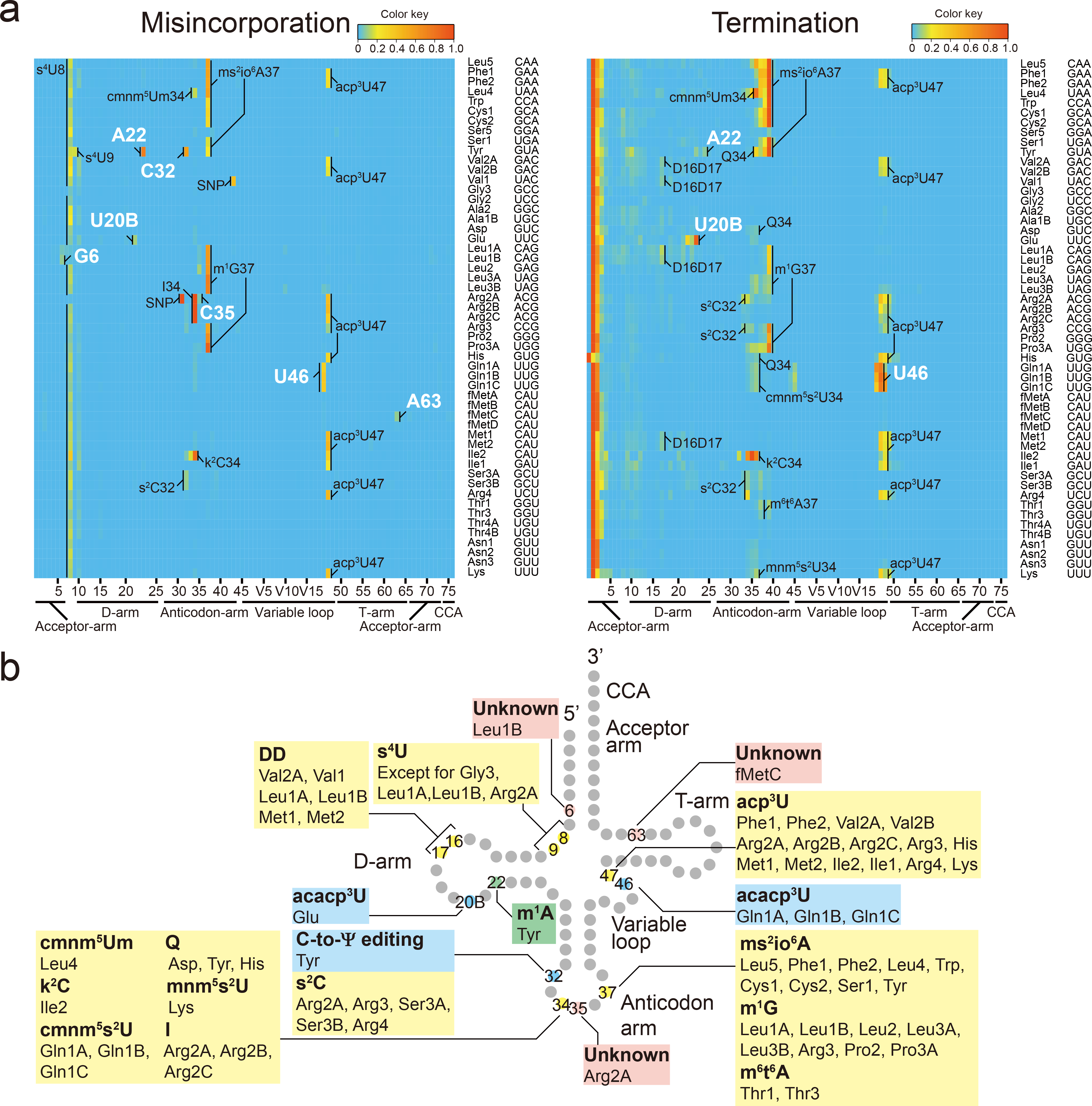
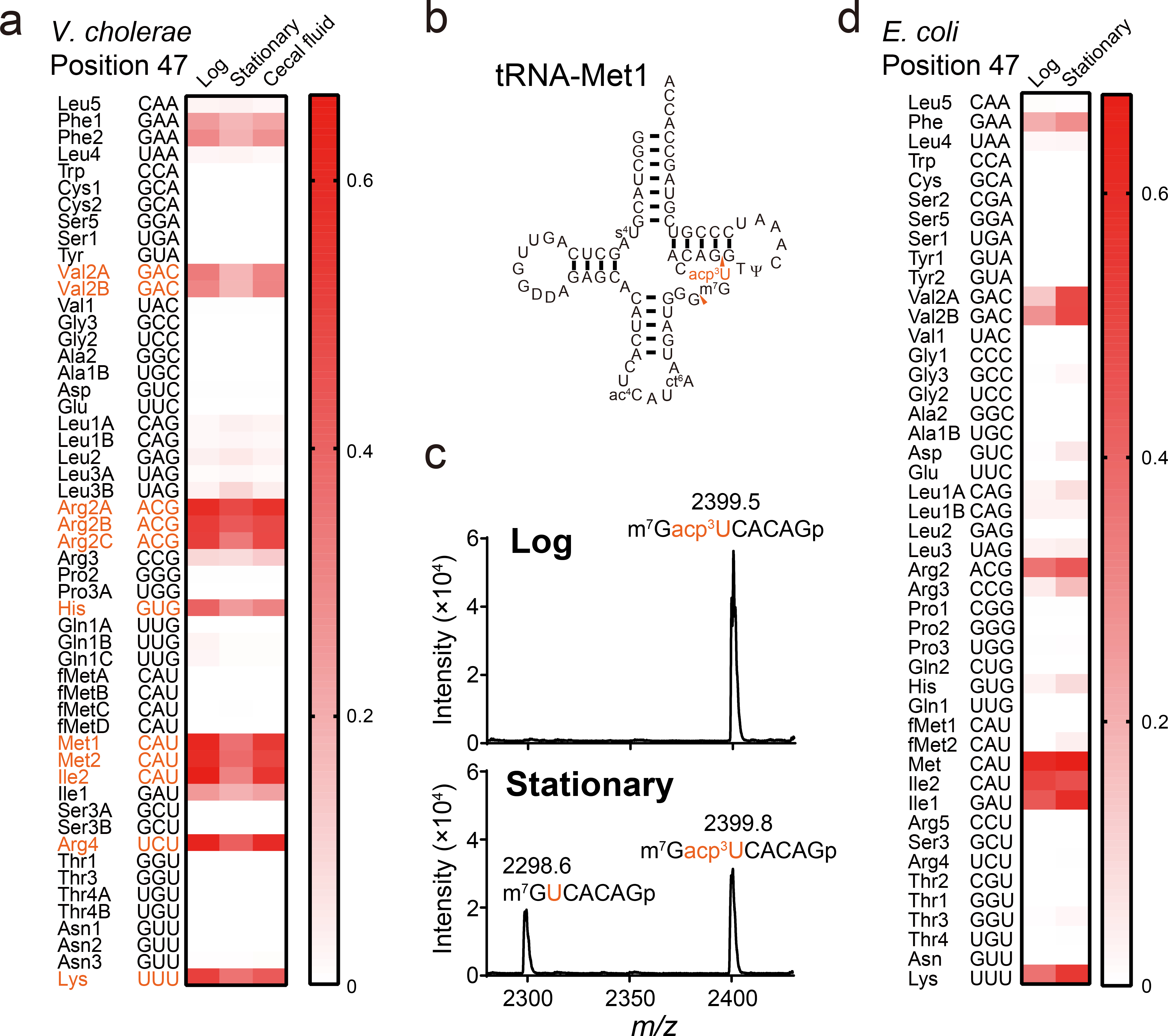
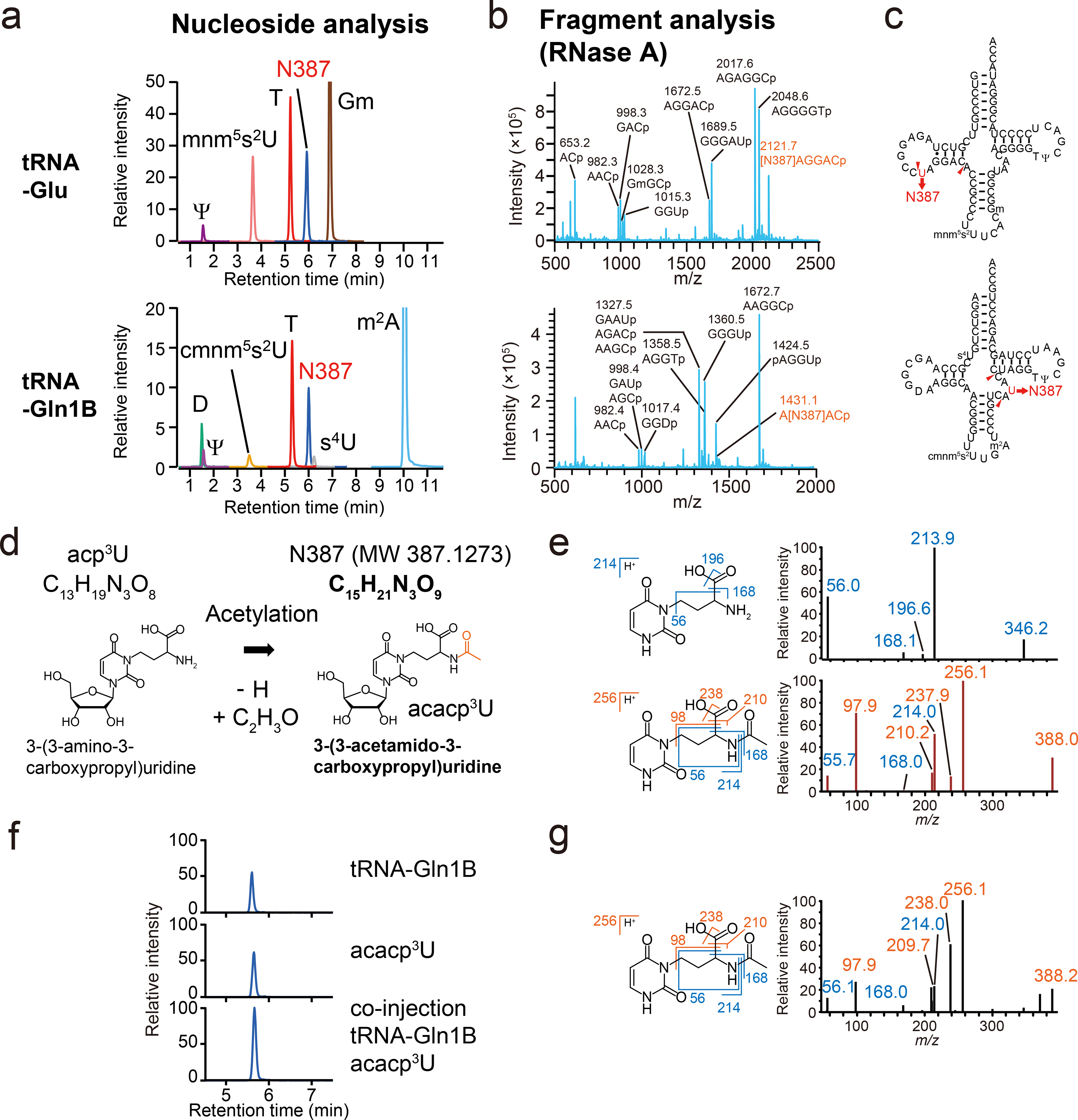
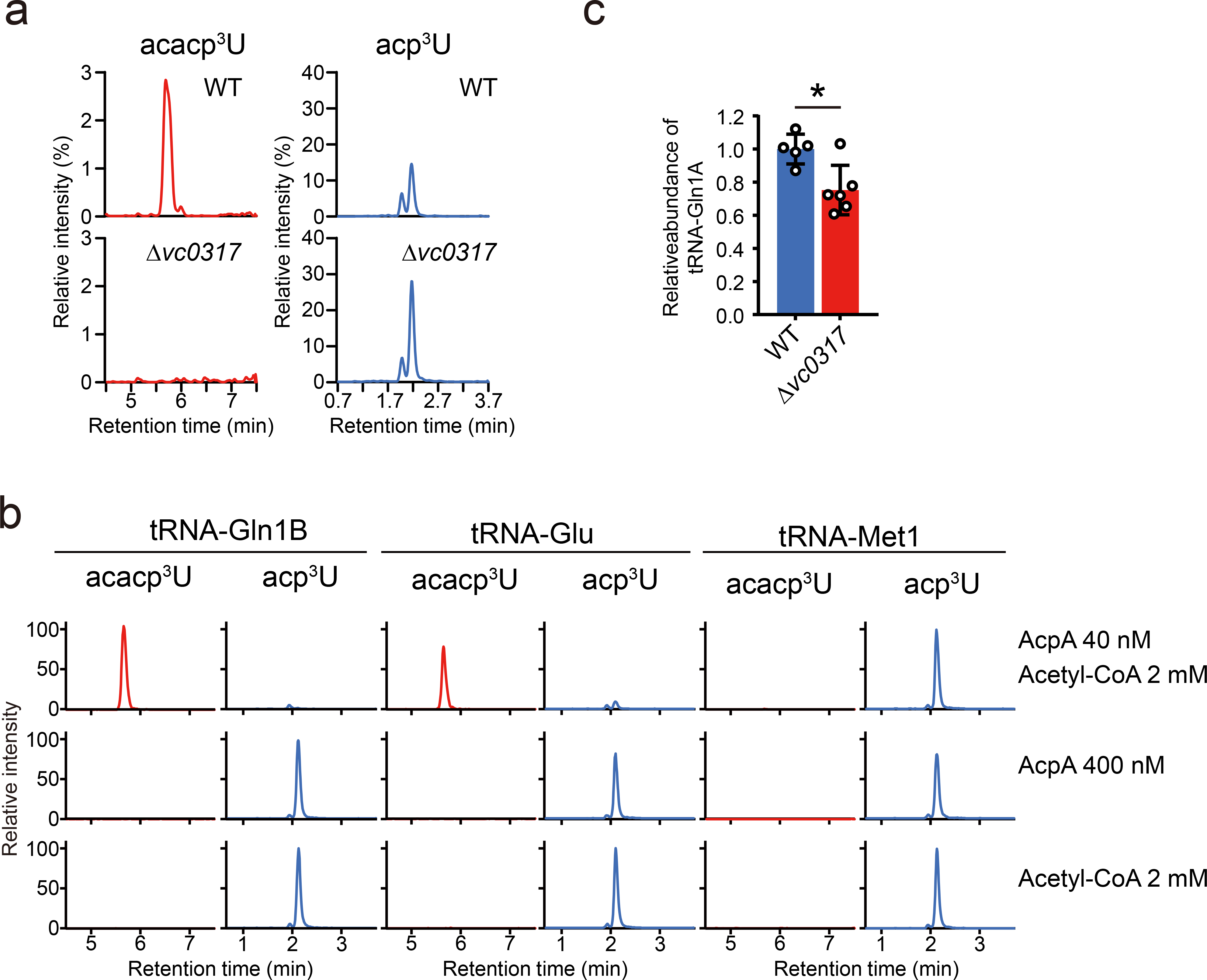
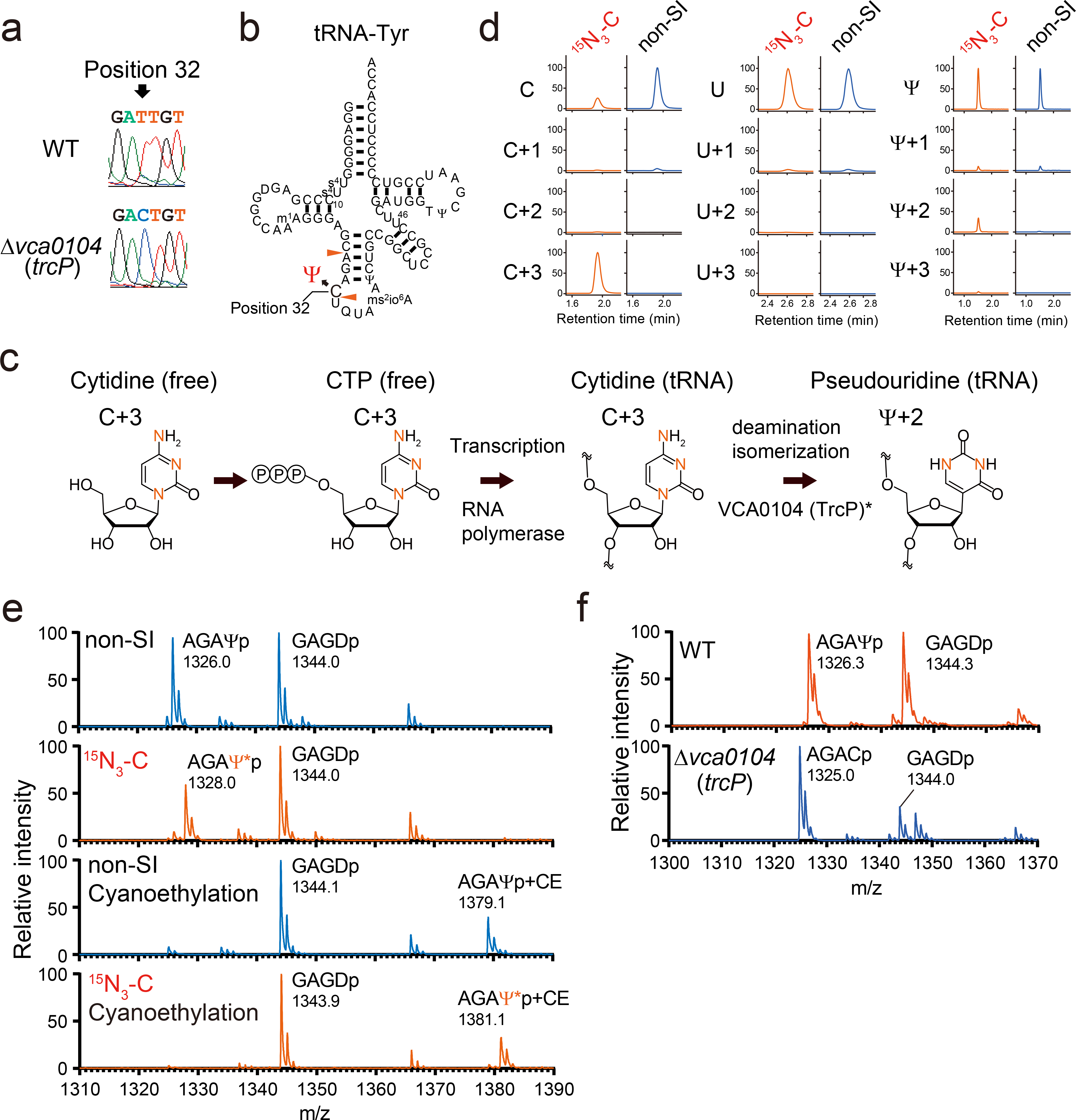
Chemical modifications of the nucleosides that comprise transfer RNAs are diverse. However, the structure, location and extent of modifications have been systematically charted in very few organisms. Here, we describe an approach in which rapid prediction of modified sites through reverse transcription-derived signatures in high-throughput transfer RNA-sequencing (tRNA-seq) data is coupled with identification of tRNA modifications through RNA mass spectrometry. Comparative tRNA-seq enabled prediction of several Vibrio cholerae modifications that are absent from Escherichia coli and also revealed the effects of various environmental conditions on V. cholerae tRNA modification. Through RNA mass spectrometric analyses, we showed that two of the V. cholerae-specific reverse transcription signatures reflected the presence of a new modification (acetylated acp3U (acacp3U)), while the other results from C-to-Ψ RNA editing, a process not described before. These findings demonstrate the utility of this approach for rapid surveillance of tRNA modification profiles and environmental control of tRNA modification.
Conflict of interest statement
Competing Financial Interests Statement
The authors declare no competing interests.
Figures
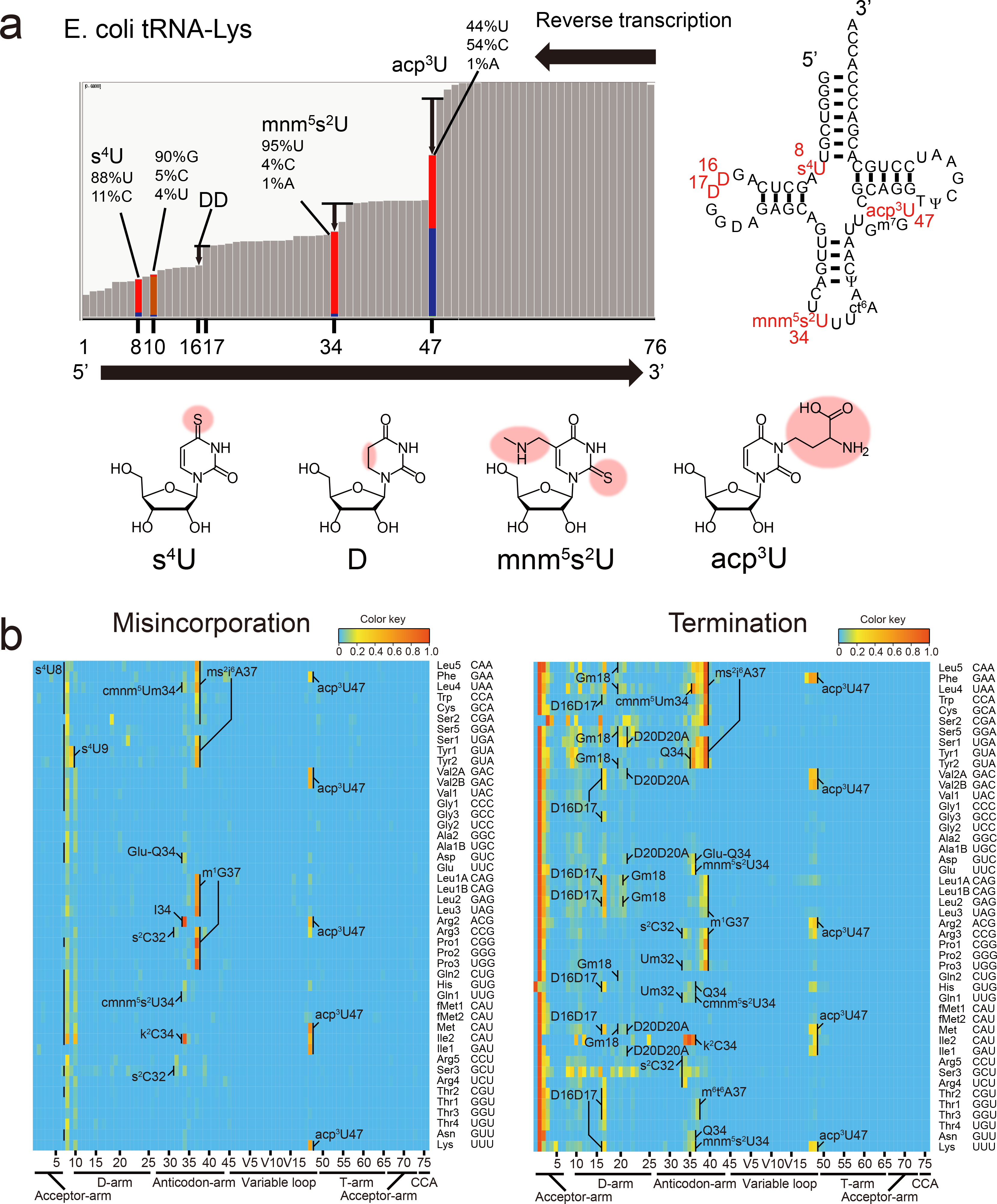
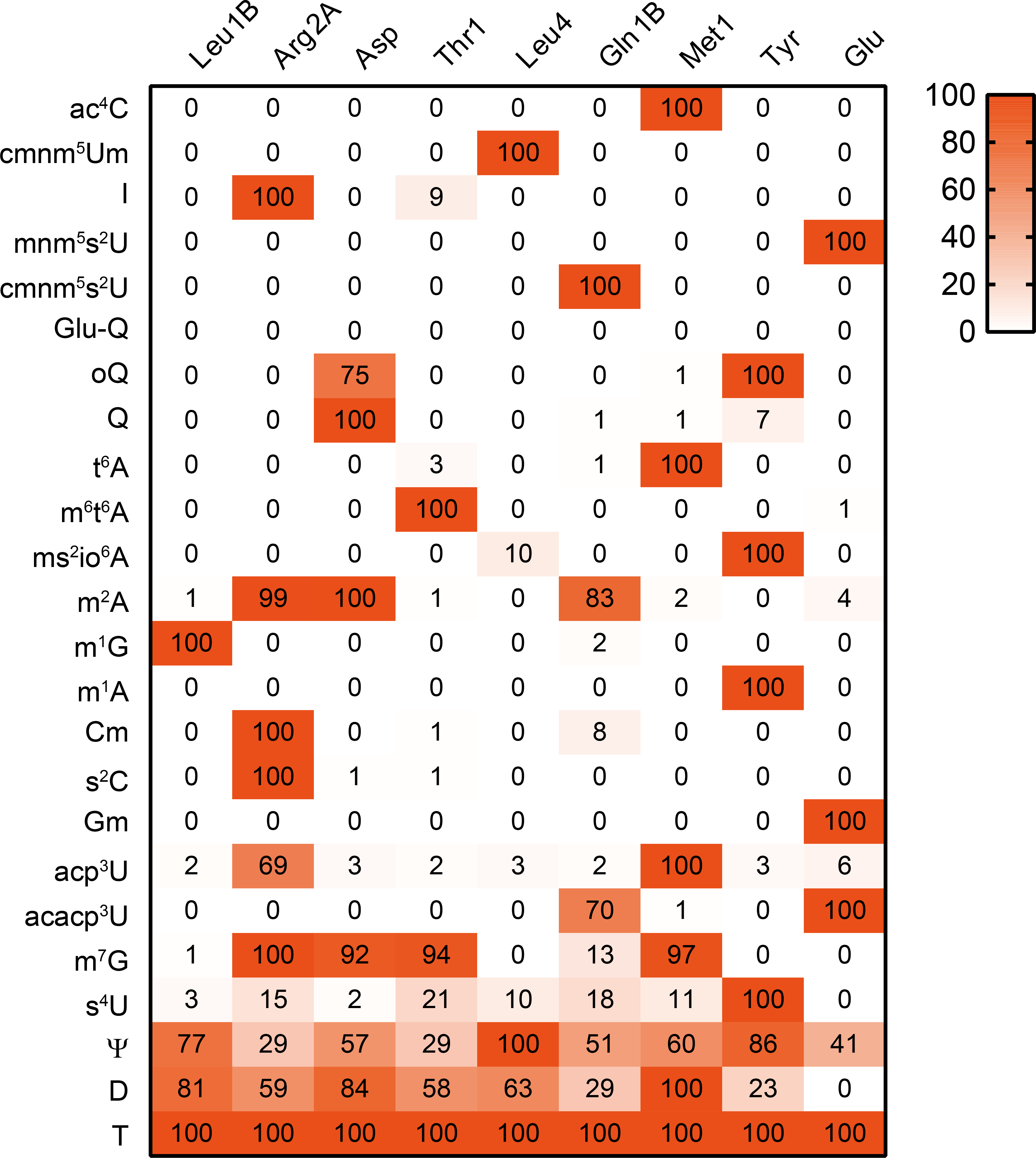
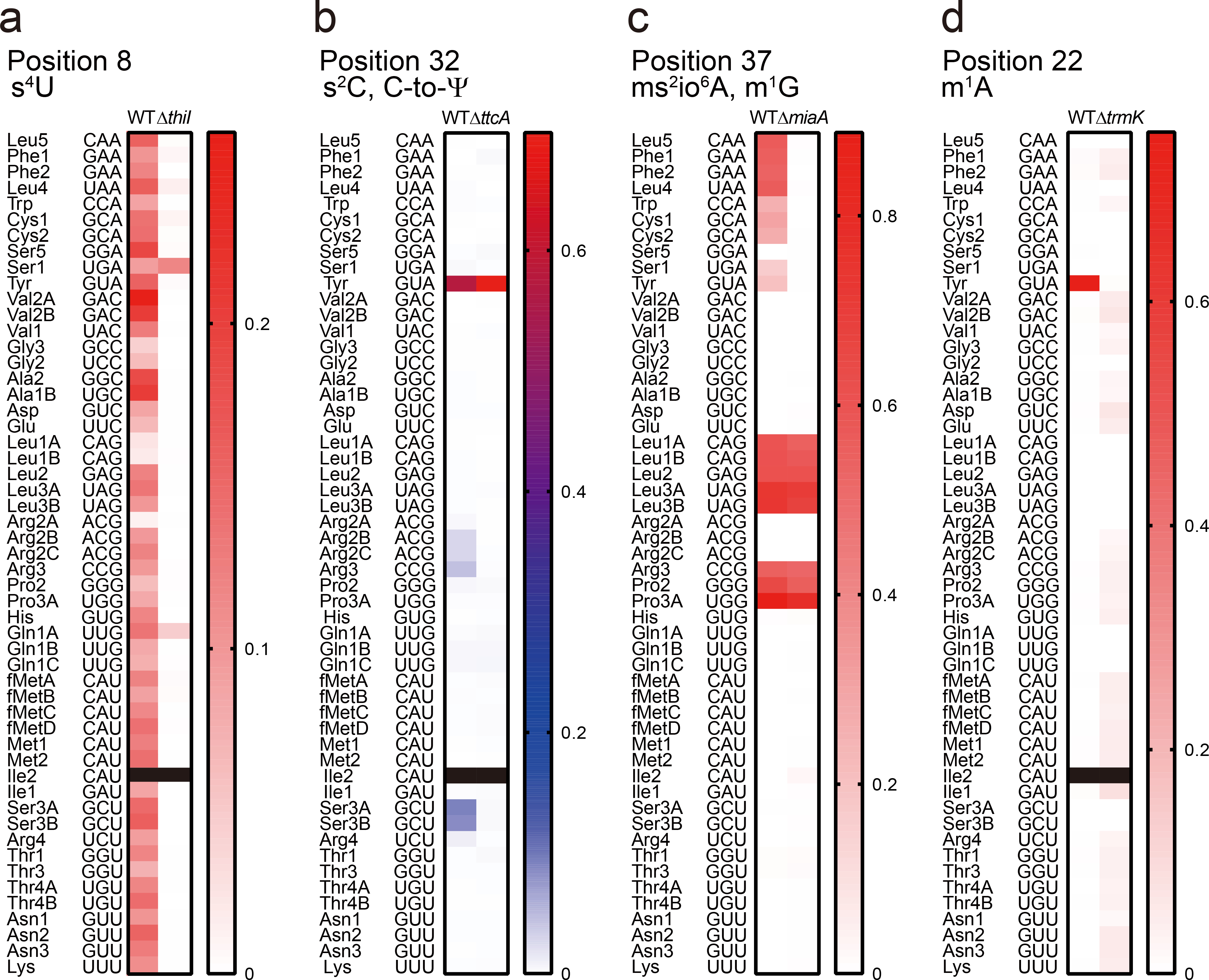
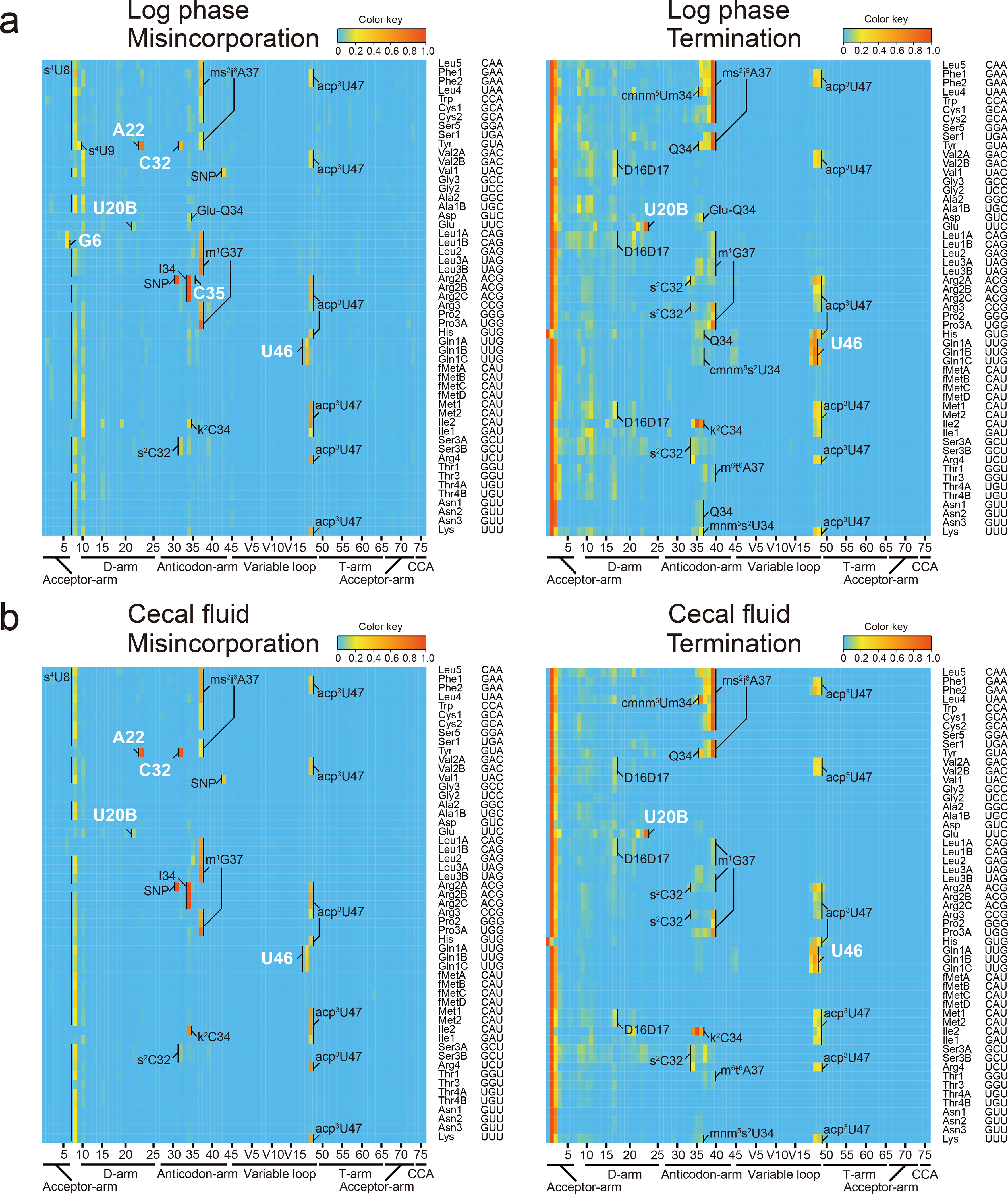
References
Methods-only references
-
- Griffey RH et al. 15N-labeled tRNA. Identification of 4-thiouridine in Escherichia coli tRNASer1 and tRNATyr2 by 1H-15N two-dimensional NMR spectroscopy. J Biol Chem 261, 12074–12078 (1986). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
