

Parthenogenesis as a Solution to Hybrid Sterility: The Mechanistic Basis of Meiotic Distortions in Clonal and Sterile Hybrids
- PMID: 32518062
- PMCID: PMC7404241
- DOI: 10.1534/genetics.119.302988
Parthenogenesis as a Solution to Hybrid Sterility: The Mechanistic Basis of Meiotic Distortions in Clonal and Sterile Hybrids
Abstract
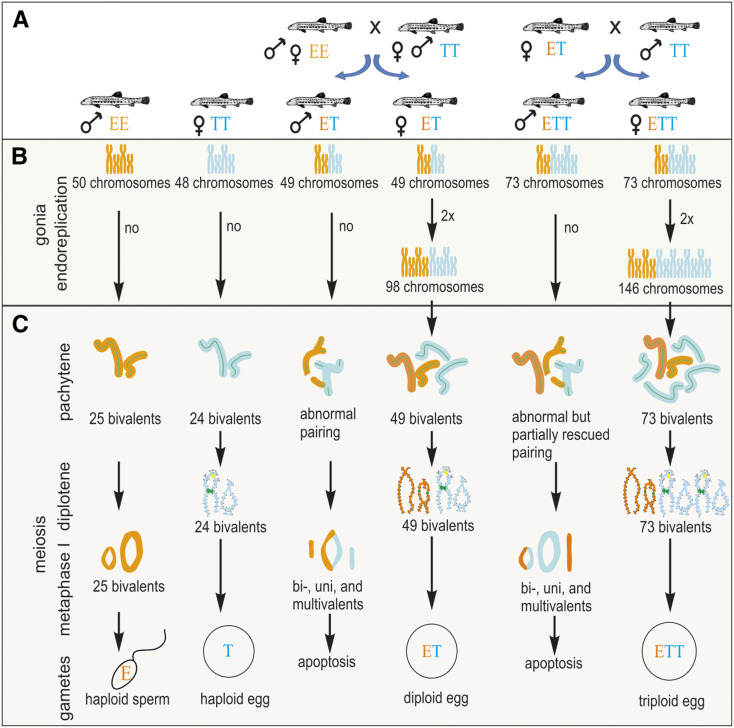
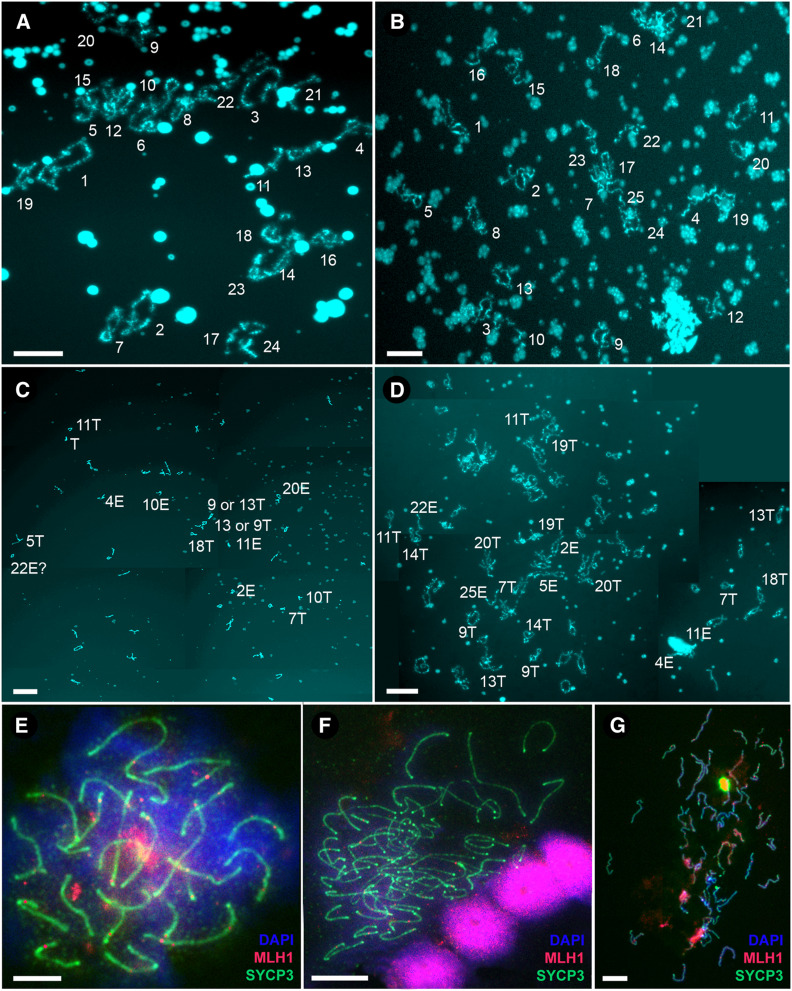
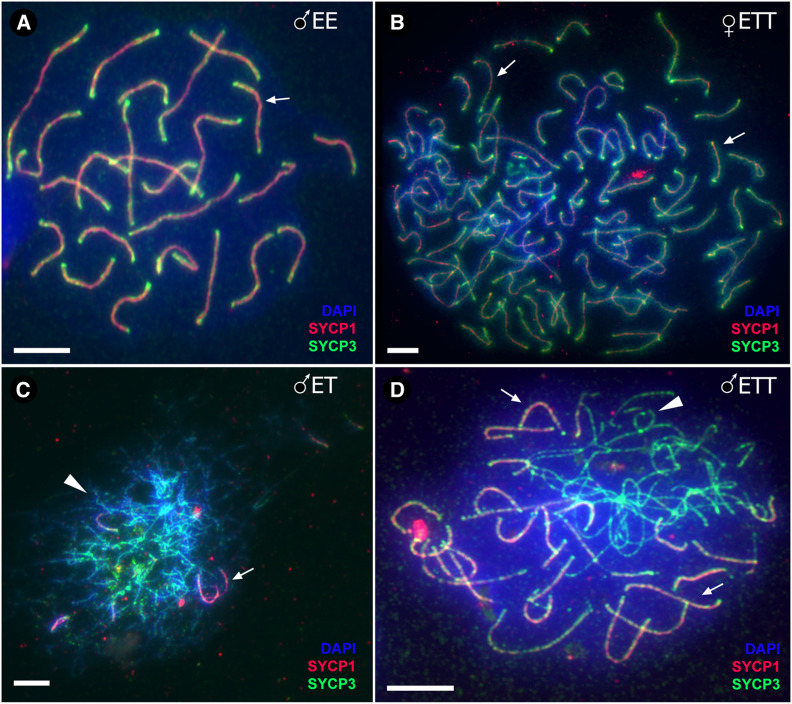
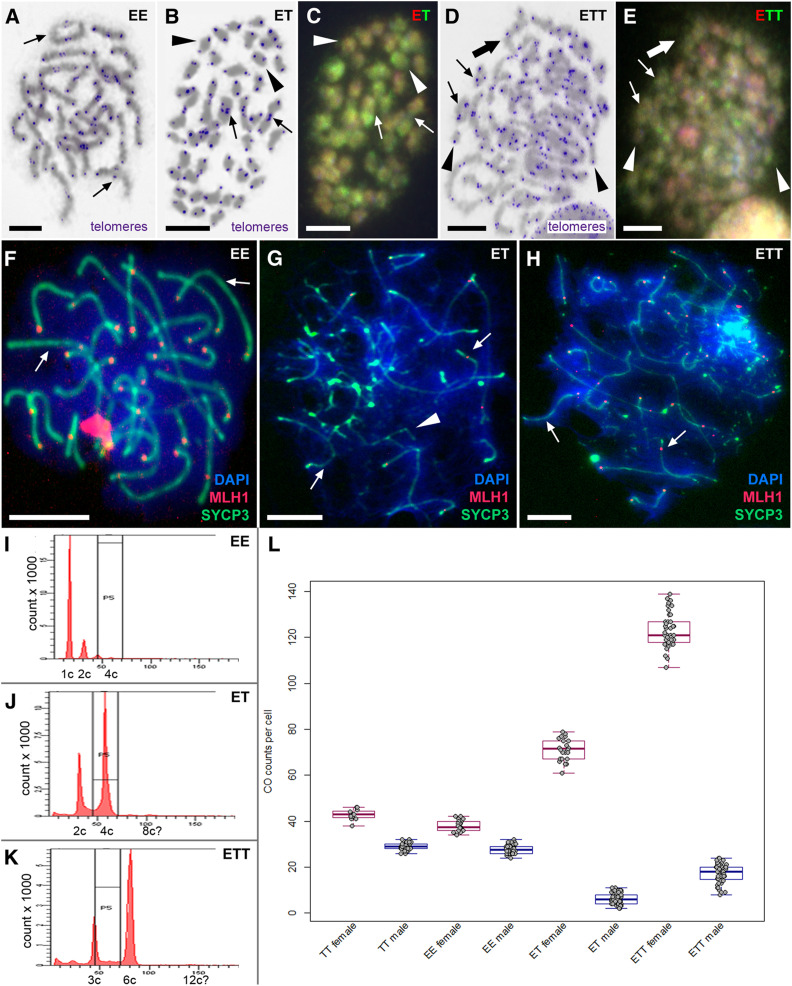
Hybrid sterility is a hallmark of speciation, but the underlying molecular mechanisms remain poorly understood. Here, we report that speciation may regularly proceed through a stage at which gene flow is completely interrupted, but hybrid sterility occurs only in male hybrids whereas female hybrids reproduce asexually. We analyzed gametogenic pathways in hybrids between the fish species Cobitis elongatoides and C. taenia, and revealed that male hybrids were sterile owing to extensive asynapsis and crossover reduction among heterospecific chromosomal pairs in their gametes, which was subsequently followed by apoptosis. We found that polyploidization allowed pairing between homologous chromosomes and therefore partially rescued the bivalent formation and crossover rates in triploid hybrid males. However, it was not sufficient to overcome sterility. In contrast, both diploid and triploid hybrid females exhibited premeiotic genome endoreplication, thereby ensuring proper bivalent formation between identical chromosomal copies. This endoreplication ultimately restored female fertility but it simultaneously resulted in the obligate production of clonal gametes, preventing any interspecific gene flow. In conclusion, we demonstrate that the emergence of asexuality can remedy hybrid sterility in a sex-specific manner and contributes to the speciation process.
Keywords: Cobitis; clonality; endoreplication; gynogenesis; hybridization; meiosis; polyploidy; speciation.
Copyright © 2020 by the Genetics Society of America.
Figures
References
-
- Avise I. J., 2008. Clonality : The genetics, Ecology, and Evolution of Sexual Abstinence in Vertebrate Animals: The Genetics, Ecology, and Evolution of Sexual Abstinence in Vertebrate Animals, Oxford University Press, Oxford: 10.1093/acprof:oso/9780195369670.001.0001 - DOI
-
- Bateson W., 1909. Heredity and variation in modern lights., pp. 85–101 in Darwin and Modern Science, Cambridge University Press, Cambridge.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
