

The J-elongated conformation of β2-glycoprotein I predominates in solution: implications for our understanding of antiphospholipid syndrome
- PMID: 32518155
- PMCID: PMC7397106
- DOI: 10.1074/jbc.RA120.013939
The J-elongated conformation of β2-glycoprotein I predominates in solution: implications for our understanding of antiphospholipid syndrome
Abstract
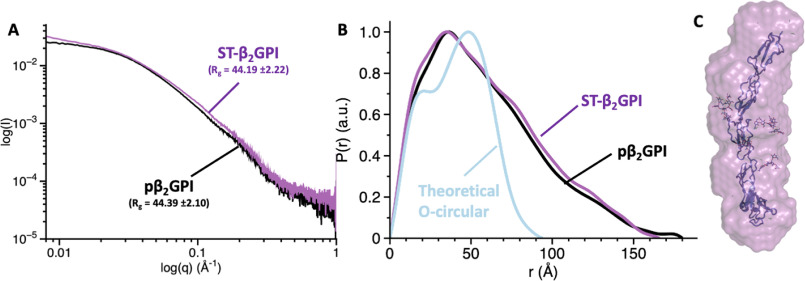
β2-Glycoprotein I (β2GPI) is an abundant plasma protein displaying phospholipid-binding properties. Because it binds phospholipids, it is a target of antiphospholipid antibodies (aPLs) in antiphospholipid syndrome (APS), a life-threatening autoimmune thrombotic disease. Indeed, aPLs prefer membrane-bound β2GPI to that in solution. β2GPI exists in two almost equally populated redox states: oxidized, in which all the disulfide bonds are formed, and reduced, in which one or more disulfide bonds are broken. Furthermore, β2GPI can adopt multiple conformations (i.e. J-elongated, S-twisted, and O-circular). While strong evidence indicates that the J-form is the structure bound to aPLs, which conformation exists and predominates in solution remains controversial, and so is the conformational pathway leading to the bound state. Here, we report that human recombinant β2GPI purified under native conditions is oxidized. Moreover, under physiological pH and salt concentrations, this oxidized form adopts a J-elongated, flexible conformation, not circular or twisted, in which the N-terminal domain I (DI) and the C-terminal domain V (DV) are exposed to the solvent. Consistent with this model, binding kinetics and mutagenesis experiments revealed that in solution the J-form interacts with negatively charged liposomes and with MBB2, a monoclonal anti-DI antibody that recapitulates most of the features of pathogenic aPLs. We conclude that the preferential binding of aPLs to phospholipid-bound β2GPI arises from the ability of its preexisting J-form to accumulate on the membranes, thereby offering an ideal environment for aPL binding. We propose that targeting the J-form of β2GPI provides a strategy to block pathogenic aPLs in APS.
Keywords: X-ray crystallography; antiphospholipid syndrome; autoimmune disease; autoimmunity; beta-2 glycoprotein I; coagulation; complement system; lipid–protein interaction; protein–protein interaction; single-molecule biophysics; structural biology; structure-function; thrombosis.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Schultze H. E., Heide K., and Haupt H. (1961) Uber ein bischer unb ekanntesniedermolekularis b2-globulins des human serums. Naturwissens-chaften 48, 719–719 10.1007/BF00620967 - DOI
-
- Forneris F., Wu J., Xue X., Ricklin D., Lin Z., Sfyroera G., Tzekou A., Volokhina E., Granneman J. C., Hauhart R., Bertram P., Liszewski M. K., Atkinson J. P., Lambris J. D., and Gros P. (2016) Regulators of complement activity mediate inhibitory mechanisms through a common C3b-binding mode. EMBO J. 35, 1133–1149 10.15252/embj.201593673 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
