

Tissue-resident memory T cell reactivation by diverse antigen-presenting cells imparts distinct functional responses
- PMID: 32525985
- PMCID: PMC7398161
- DOI: 10.1084/jem.20192291
Tissue-resident memory T cell reactivation by diverse antigen-presenting cells imparts distinct functional responses
Abstract
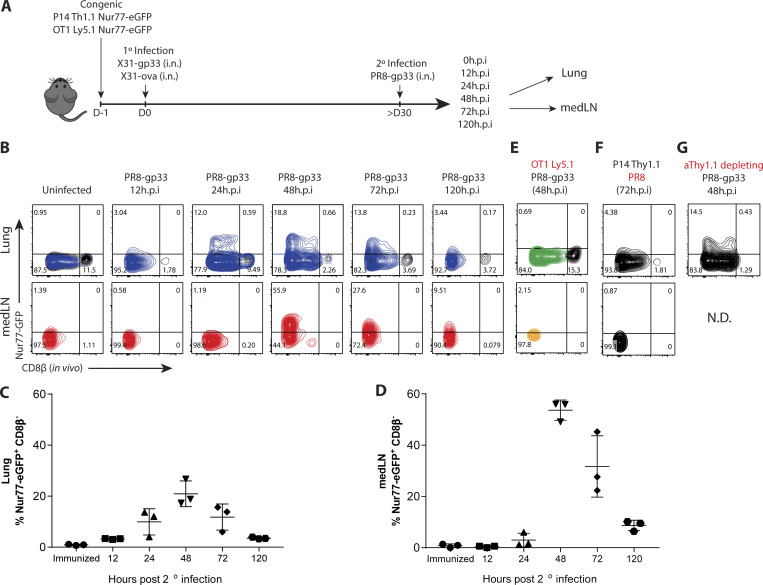
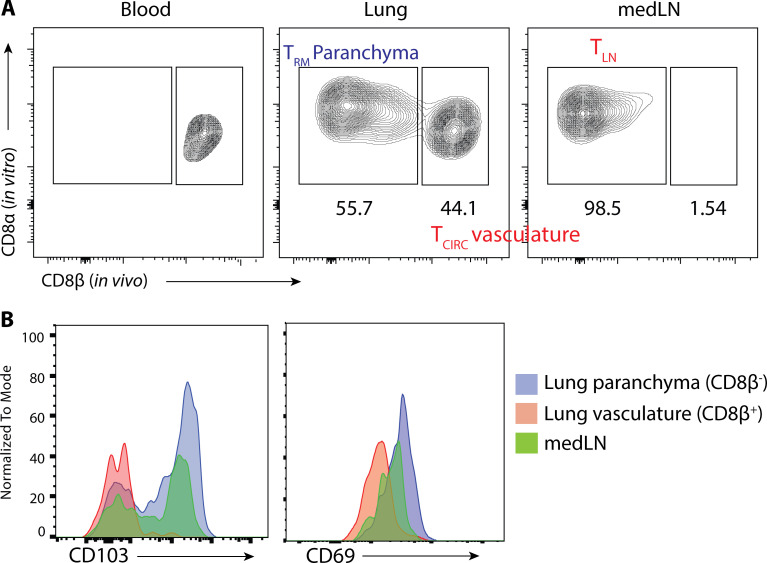
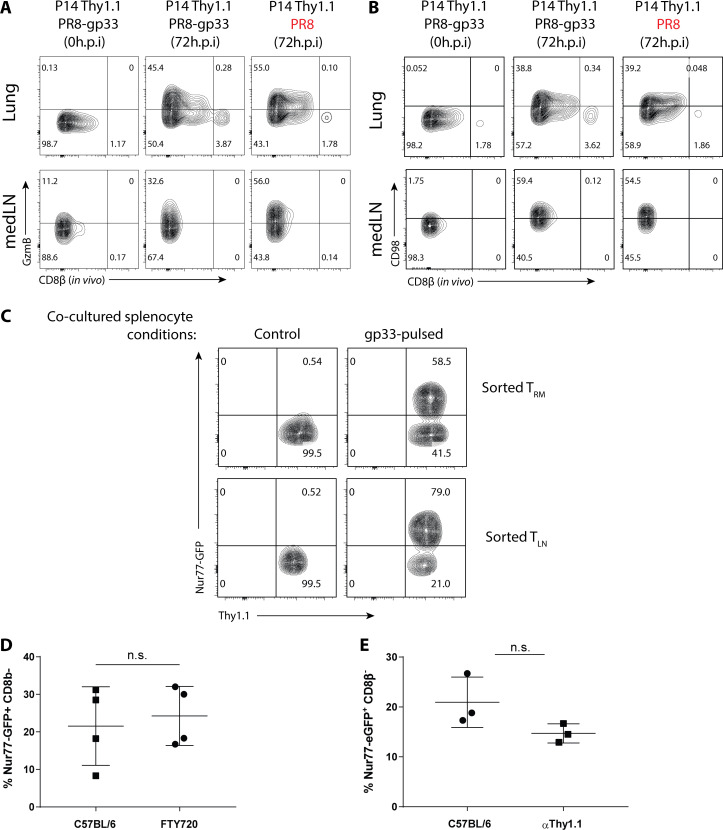
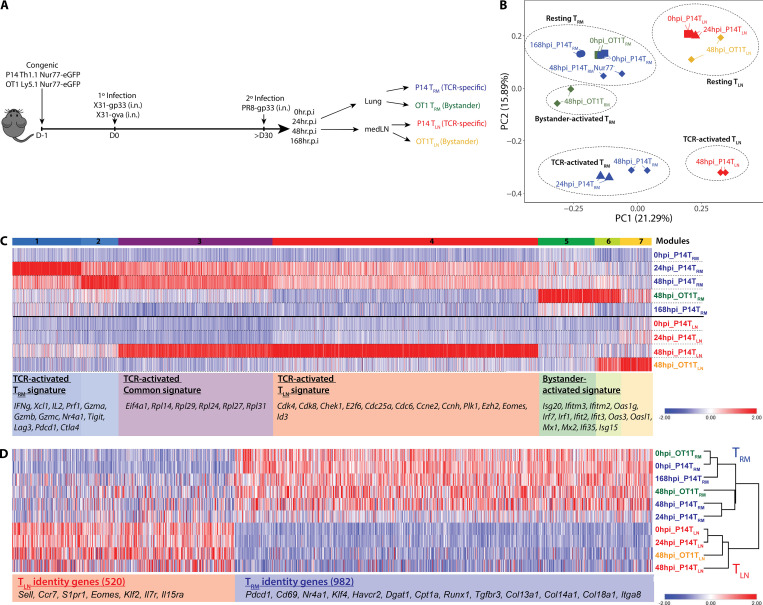
CD8+ tissue-resident memory T cells (TRM cells) are poised at the portals of infection and provide long-term protective immunity. Despite their critical roles, the precise mechanics governing TRM cell reactivation in situ are unknown. Using a TCR-transgenic Nur77-GFP reporter to distinguish "antigen-specific" from "bystander" reactivation, we demonstrate that lung CD8+ TRM cells are reactivated more quickly, yet less efficiently, than their counterparts in the draining LNs (TLN cells). Global profiling of reactivated memory T cells revealed tissue-defined and temporally regulated recall response programs. Unlike the reactivation of CD8+ TLN cells, which is strictly dependent on CD11c+XCR1+ APCs, numerous antigen-presenting partners, both hematopoietic and non-hematopoietic, were sufficient to reactivate lung CD8+ TRM cells, but the quality of TRM cell functional responses depended on the identity of the APCs. Together, this work uncovers fundamental differences in the activation kinetics, mechanics, and effector responses between CD8+ memory T cells in peripheral vs. lymphoid organs, revealing a novel tissue-specific paradigm for the reactivation of memory CD8+ T cells.
© 2020 Low et al.
Conflict of interest statement
Disclosures: S.M. Kaech reported personal fees from Celsius Therapeutics and personal fees from the JEM Editorial Board outside the submitted work. No other disclosures were reported.
Figures

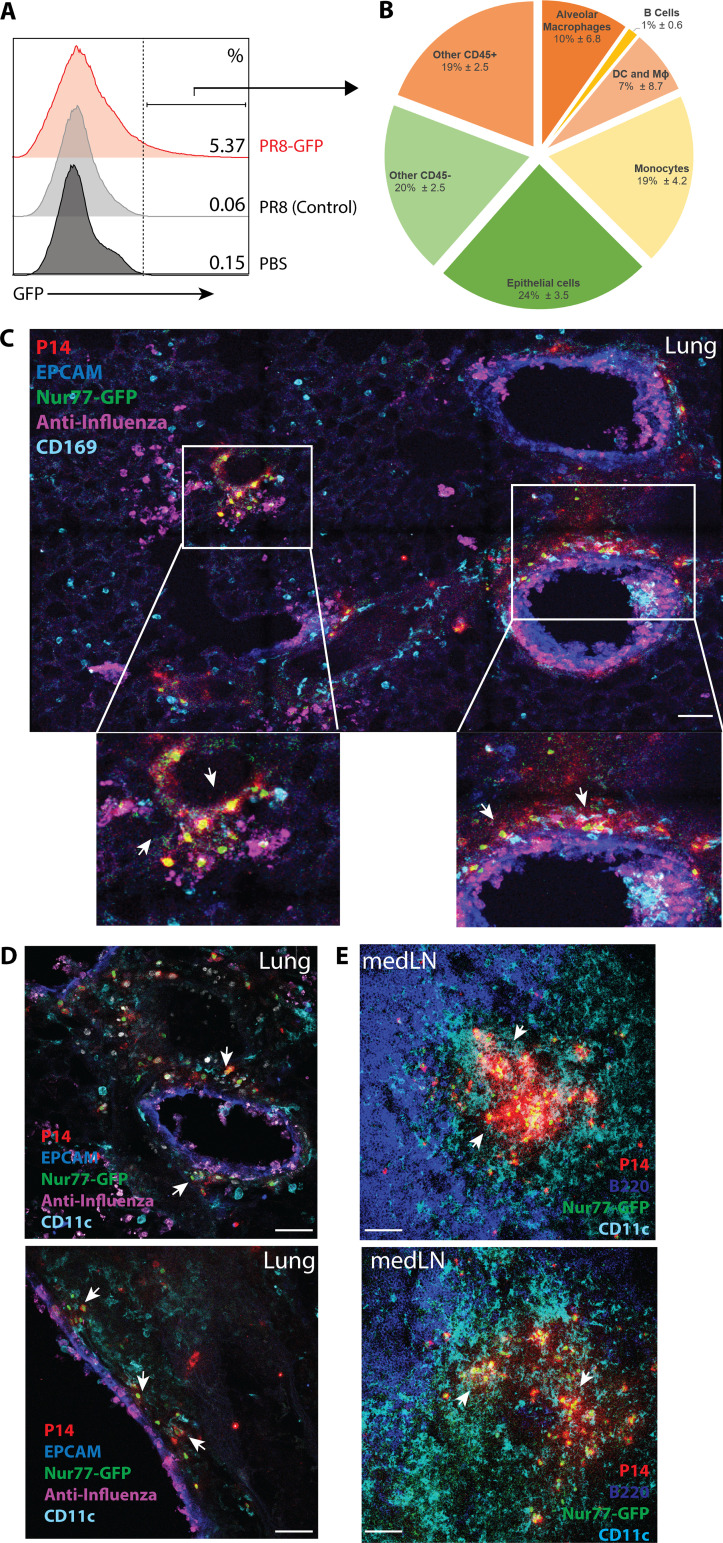


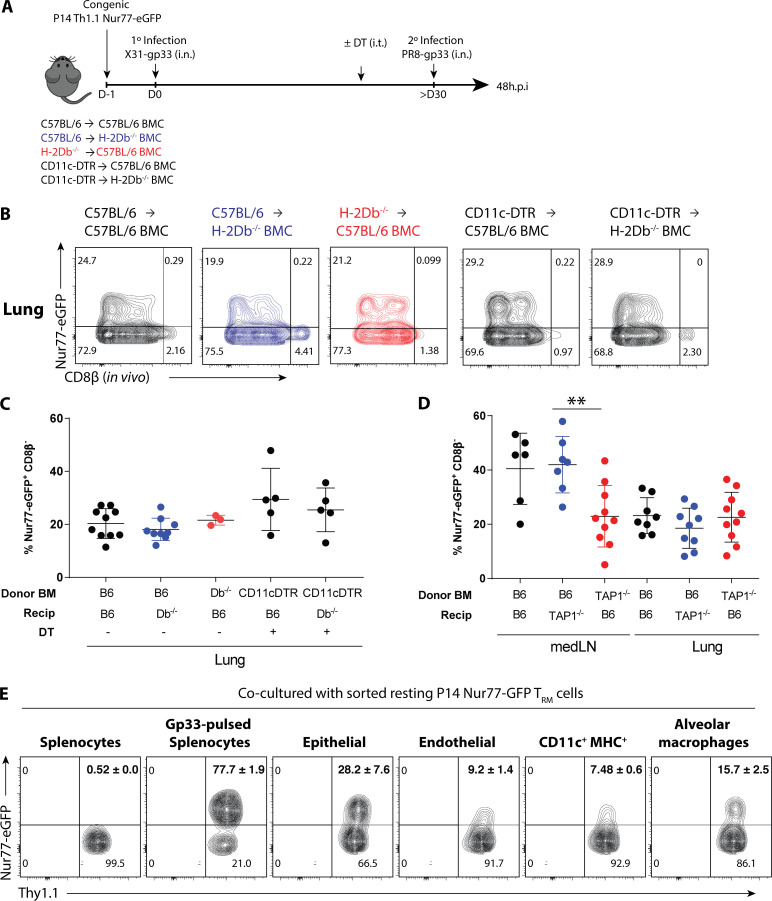
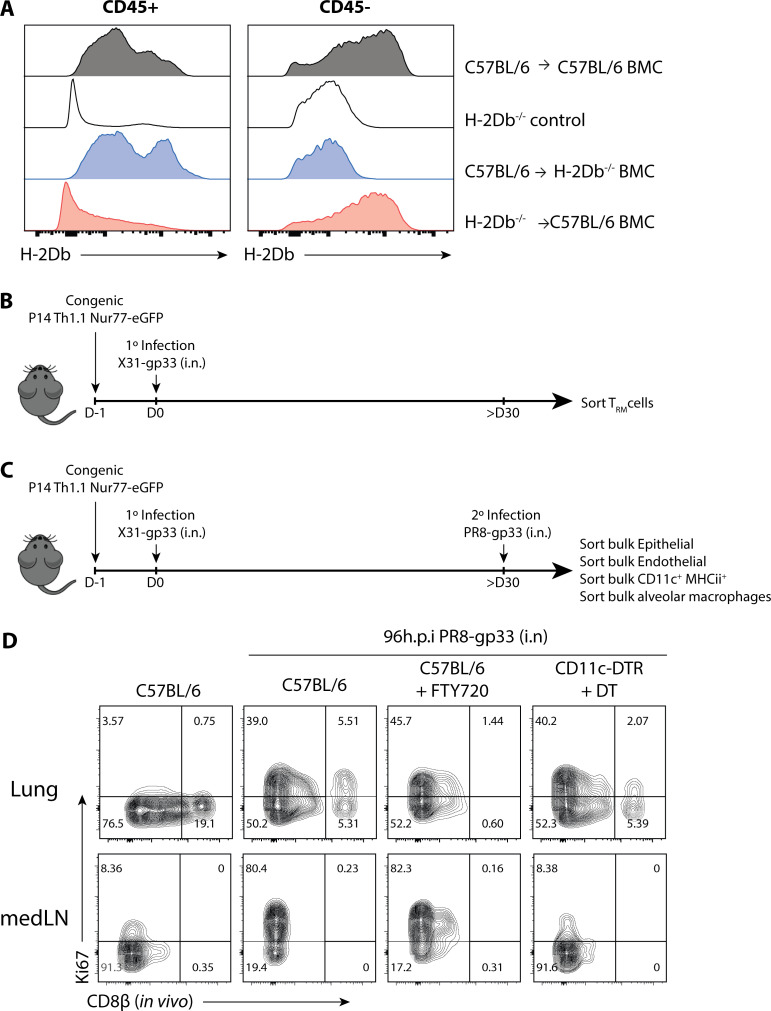
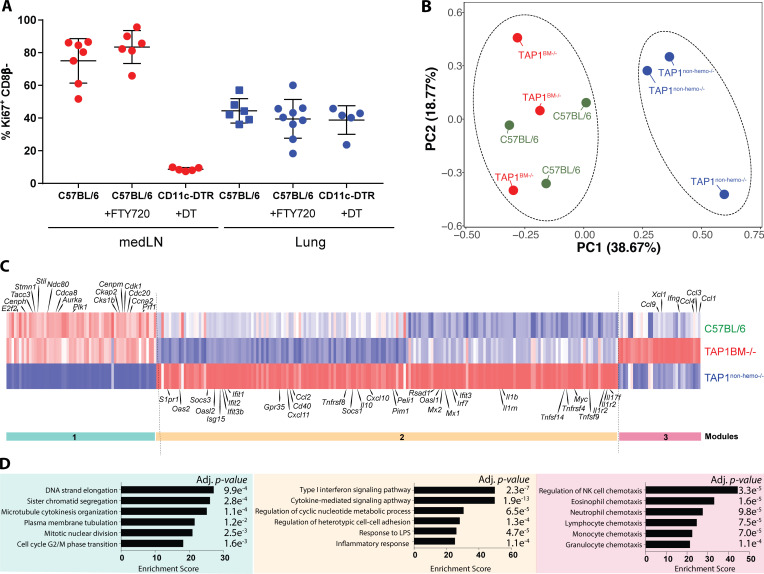
References
-
- De Baets S., Verhelst J., Van den Hoecke S., Smet A., Schotsaert M., Job E.R., Roose K., Schepens B., Fiers W., and Saelens X.. 2015. A GFP expressing influenza A virus to report in vivo tropism and protection by a matrix protein 2 ectodomain-specific monoclonal antibody. PLoS One. 10 e0121491 10.1371/journal.pone.0121491 - DOI - PMC - PubMed
-
- Beura L.K., Mitchell J.S., Thompson E.A., Schenkel J.M., Mohammed J., Wijeyesinghe S., Fonseca R., Burbach B.J., Hickman H.D., Vezys V., et al. . 2018a Intravital mucosal imaging of CD8+ resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory. Nat. Immunol. 19:173–182. 10.1038/s41590-017-0029-3 - DOI - PMC - PubMed
-
- Beura L.K., Wijeyesinghe S., Thompson E.A., Macchietto M.G., Rosato P.C., Pierson M.J., Schenkel J.M., Mitchell J.S., Vezys V., Fife B.T., et al. . 2018b T Cells in Nonlymphoid Tissues Give Rise to Lymph-Node-Resident Memory T Cells. Immunity. 48:327–338.e5. 10.1016/j.immuni.2018.01.015 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
