

Membrane-proximal F-actin restricts local membrane protrusions and directs cell migration
- PMID: 32527825
- PMCID: PMC8283920
- DOI: 10.1126/science.aay7794
Membrane-proximal F-actin restricts local membrane protrusions and directs cell migration
Abstract
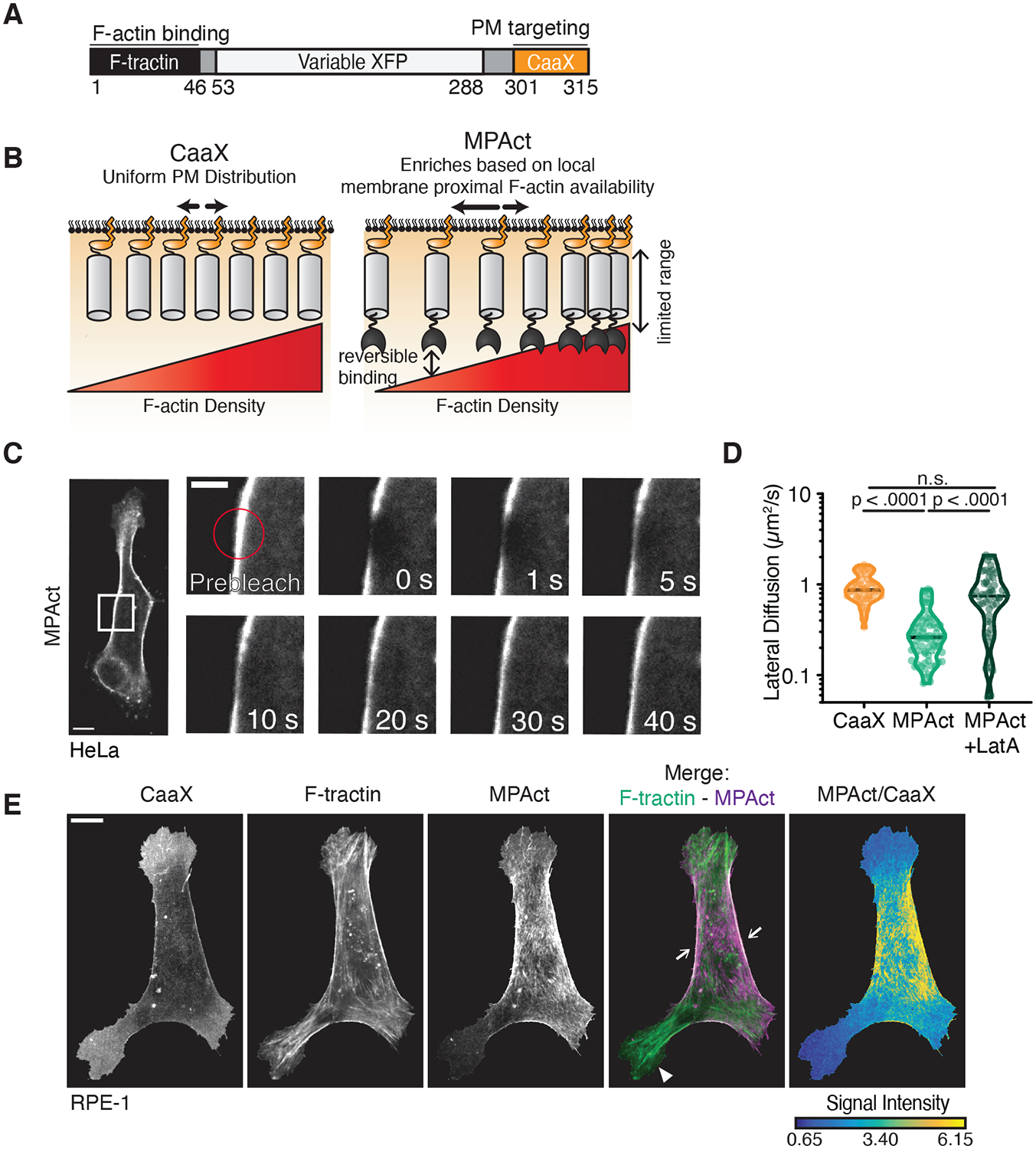
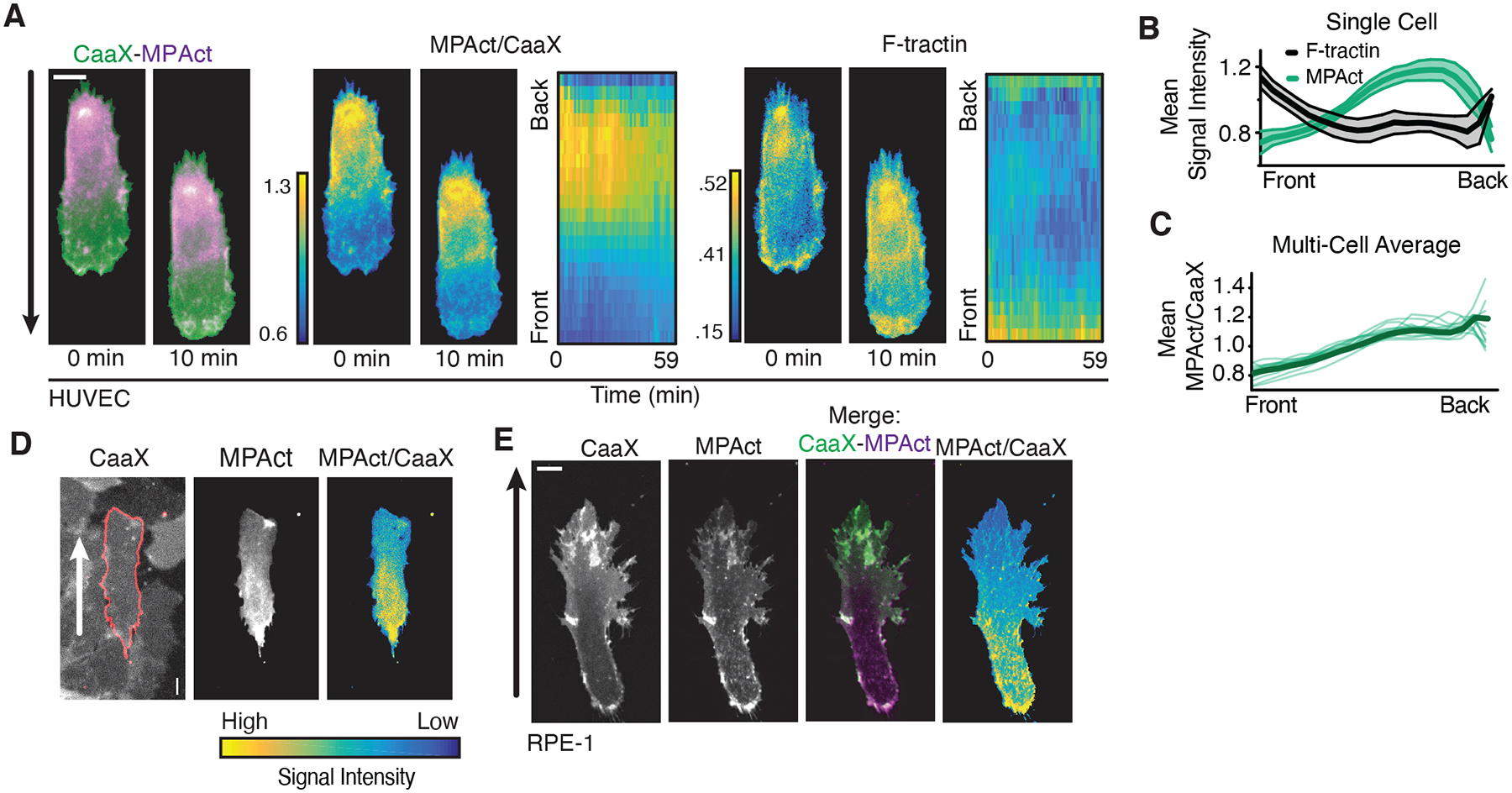
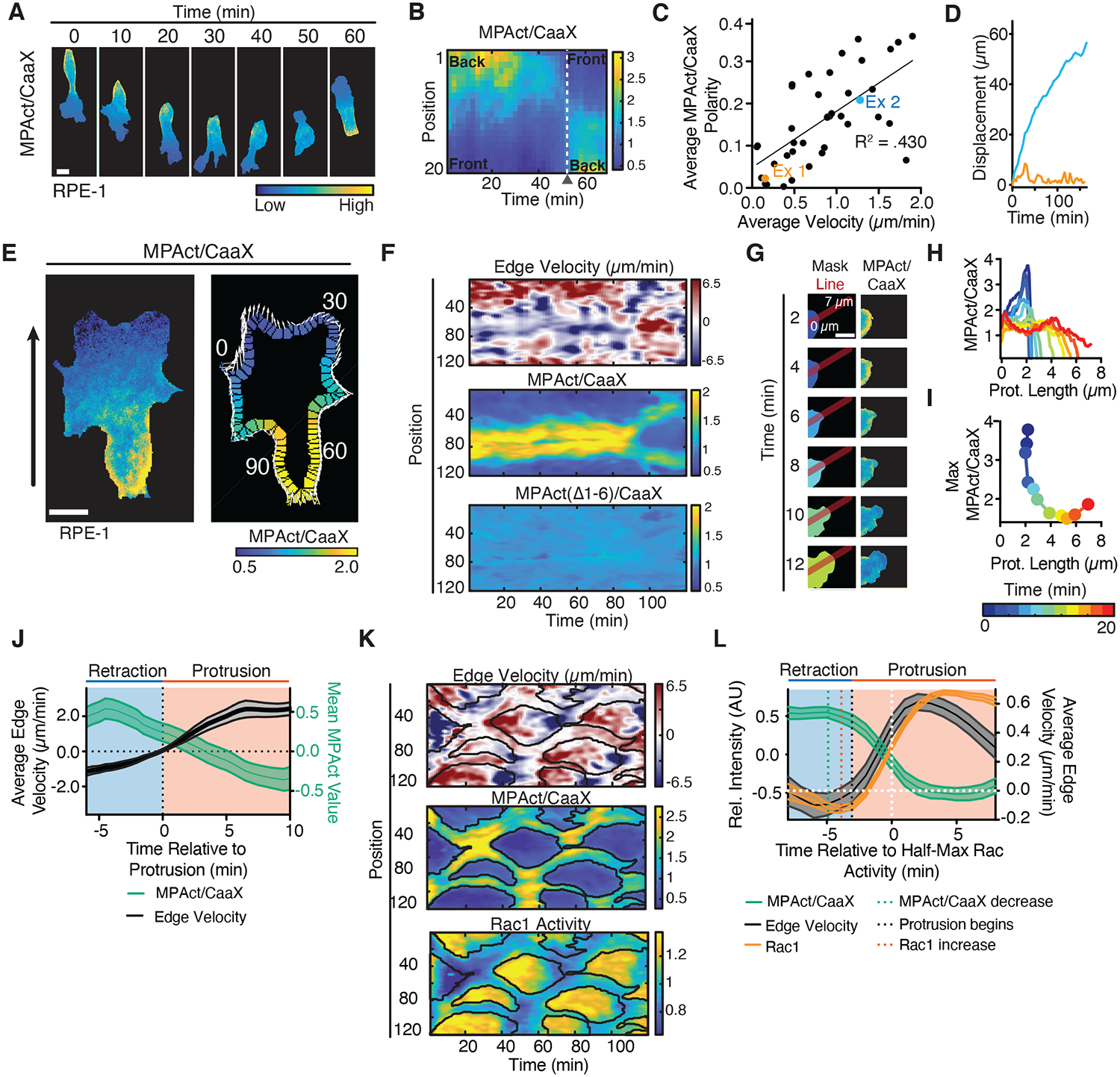
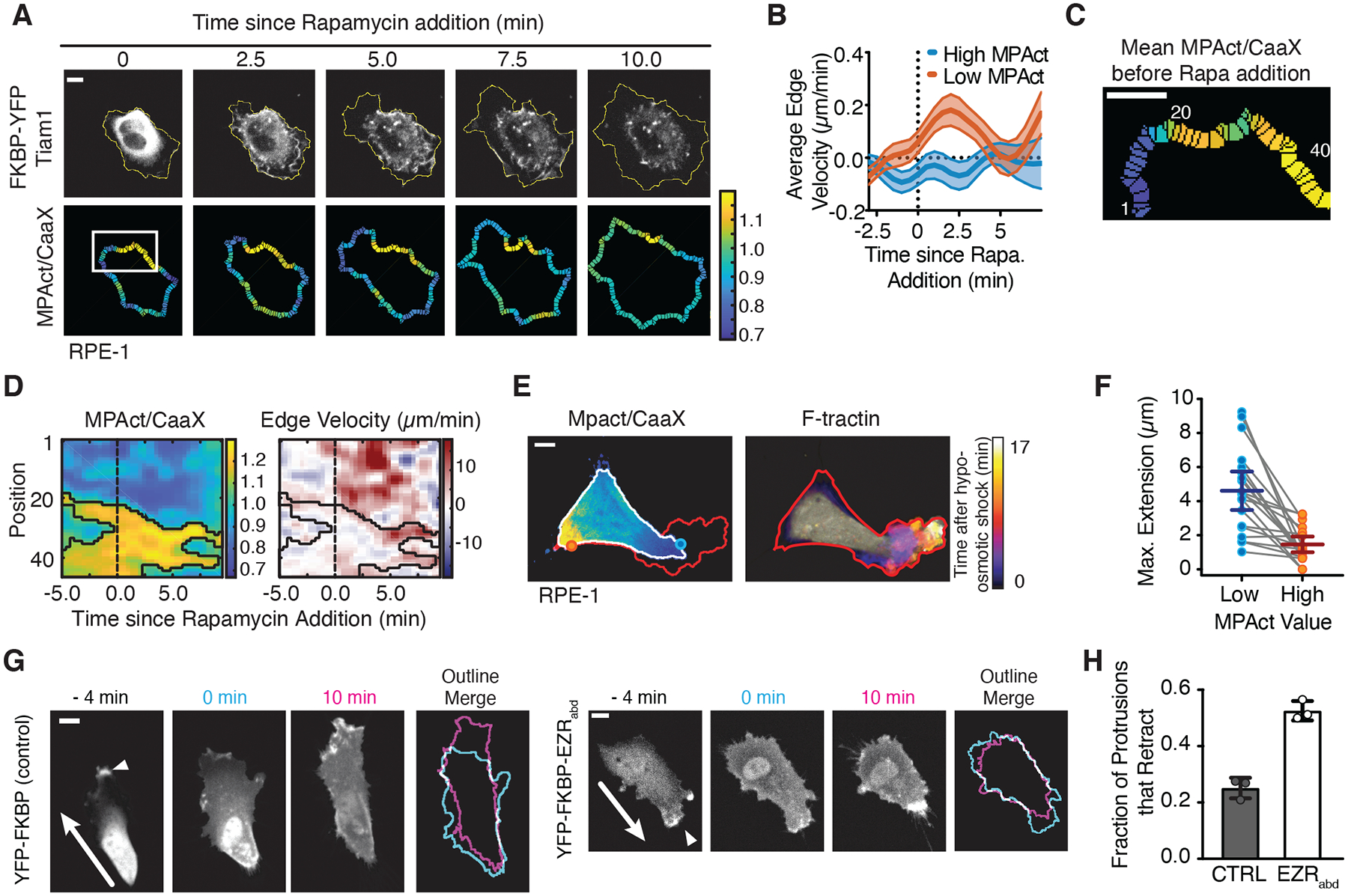
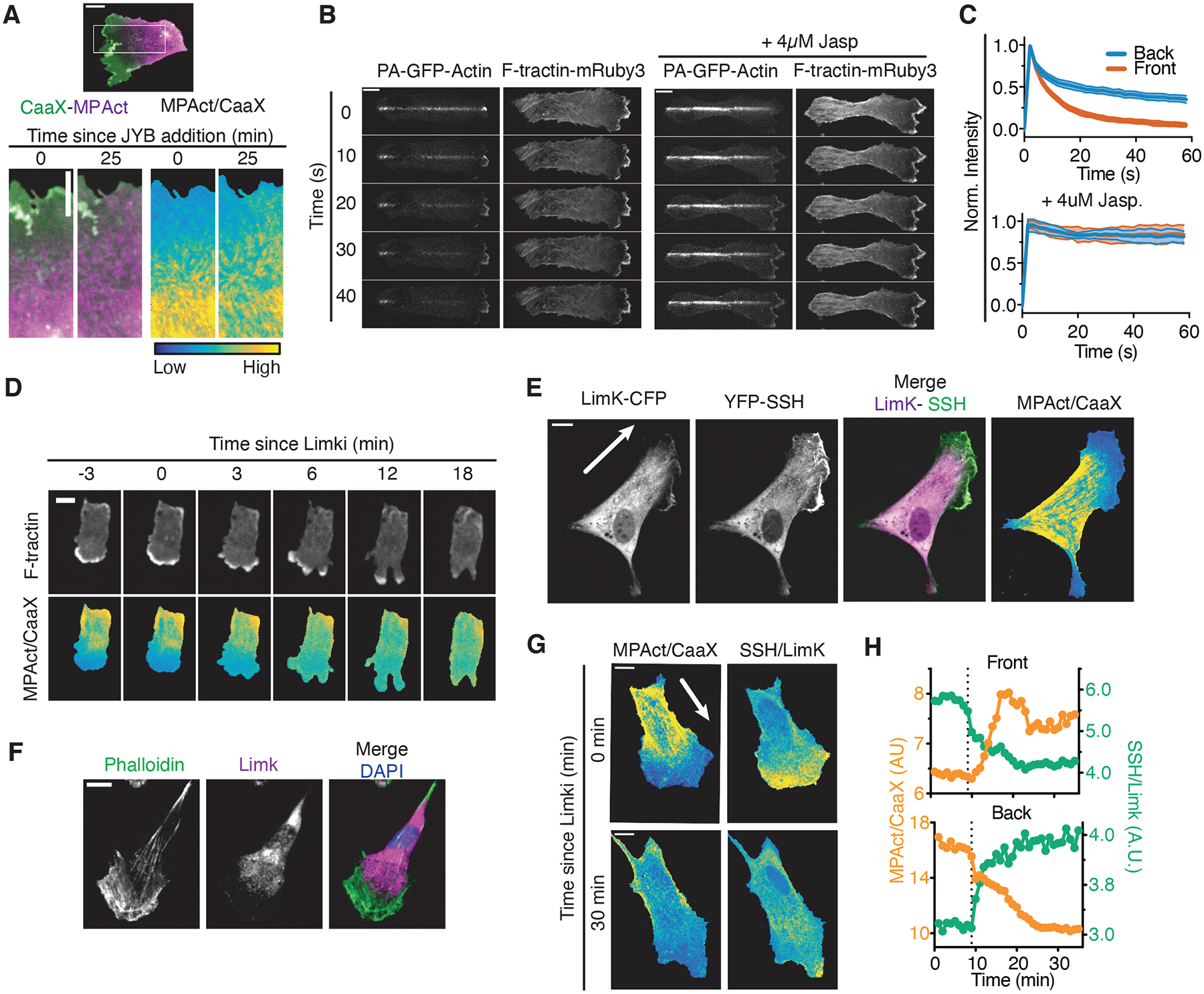
Cell migration is driven by local membrane protrusion through directed polymerization of F-actin at the front. However, F-actin next to the plasma membrane also tethers the membrane and thus resists outgoing protrusions. Here, we developed a fluorescent reporter to monitor changes in the density of membrane-proximal F-actin (MPA) during membrane protrusion and cell migration. Unlike the total F-actin concentration, which was high in the front of migrating cells, MPA density was low in the front and high in the back. Back-to-front MPA density gradients were controlled by higher cofilin-mediated turnover of F-actin in the front. Furthermore, nascent membrane protrusions selectively extended outward from areas where MPA density was reduced. Thus, locally low MPA density directs local membrane protrusions and stabilizes cell polarization during cell migration.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Salbreux G, Charras G, Paluch E, Actin cortex mechanics and cellular morphogenesis. Trends Cell Biol 22, 536–545 (2012). - PubMed
-
- Chalut KJ, Paluch EK, The Actin Cortex: A Bridge between Cell Shape and Function. Developmental Cell 38, 571–573 (2016). - PubMed
-
- Ostrowski PP, Grinstein S, Freeman SA, Diffusion Barriers, Mechanical Forces, and the Biophysics of Phagocytosis. Developmental Cell 38, 135–146 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
