

Autophagy-mediated apoptosis eliminates aneuploid cells in a mouse model of chromosome mosaicism
- PMID: 32528010
- PMCID: PMC7290028
- DOI: 10.1038/s41467-020-16796-3
Autophagy-mediated apoptosis eliminates aneuploid cells in a mouse model of chromosome mosaicism
Abstract
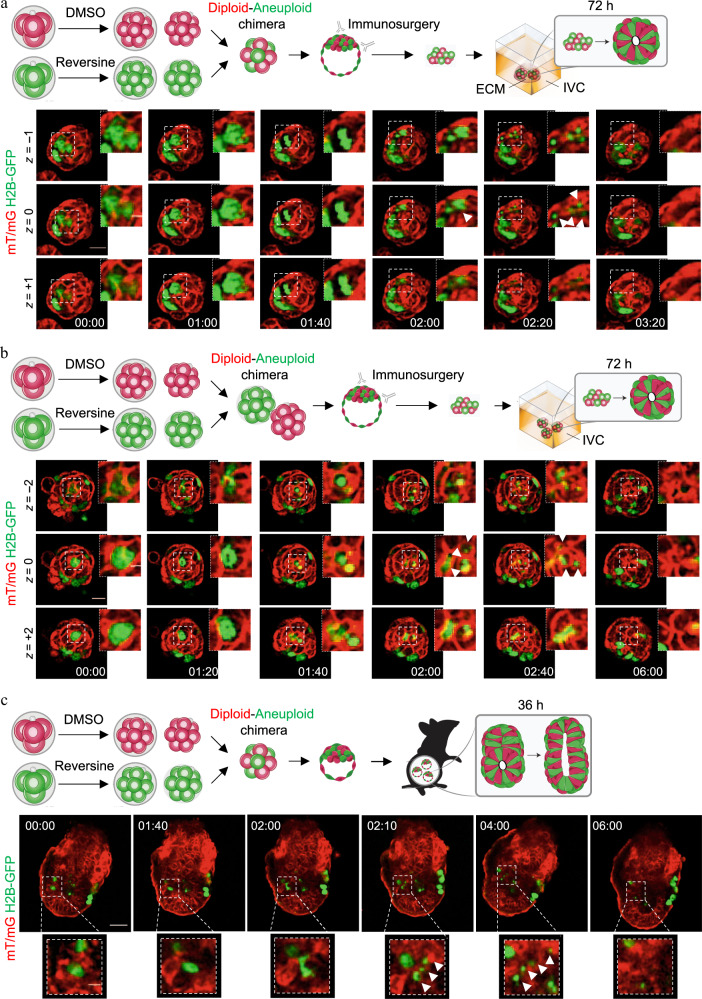
The high incidence of aneuploidy in the embryo is considered the principal cause for low human fecundity. However, the prevalence of aneuploidy dramatically declines as pregnancy progresses, with the steepest drop occurring as the embryo completes implantation. Despite the fact that the plasticity of the embryo in dealing with aneuploidy is fundamental to normal development, the mechanisms responsible for eliminating aneuploid cells are unclear. Here, using a mouse model of chromosome mosaicism, we show that aneuploid cells are preferentially eliminated from the embryonic lineage in a p53-dependent process involving both autophagy and apoptosis before, during and after implantation. Moreover, we show that diploid cells in mosaic embryos undertake compensatory proliferation during the implantation stages to confer embryonic viability. Together, our results indicate a close link between aneuploidy, autophagy, and apoptosis to refine the embryonic cell population and ensure only chromosomally fit cells proceed through development of the fetus.
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- Delhanty JDA. Preimplantation genetics: an explanation for poor human fertility? Ann. Hum. Genet. 2001;65:331–338. - PubMed
-
- Macklon NS, Geraedts JPM, Fauser BCJM. Conception to ongoing pregnancy: the ‘black box’ of early pregnancy loss. Hum. Reprod. Update. 2002;8:333–343. - PubMed
-
- Magli MC, et al. Chromosome mosaicism in day 3 aneuploid embryos that develop to morphologically normal blastocysts in vitro. Hum. Reprod. 2000;15:1781–1786. - PubMed
-
- Rubio C, et al. Impact of chromosomal abnormalities on preimplantation embryo development. Prenat. Diagn. 2007;27:748–756. - PubMed
-
- Van Echten-Arends J, et al. Chromosomal mosaicism in human pre-implantation embryos: a systematic review. Hum. Reprod. Update. 2011;17:620–627. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
