

Bacterial symbionts support larval sap feeding and adult folivory in (semi-)aquatic reed beetles
- PMID: 32528063
- PMCID: PMC7289800
- DOI: 10.1038/s41467-020-16687-7
Bacterial symbionts support larval sap feeding and adult folivory in (semi-)aquatic reed beetles
Abstract
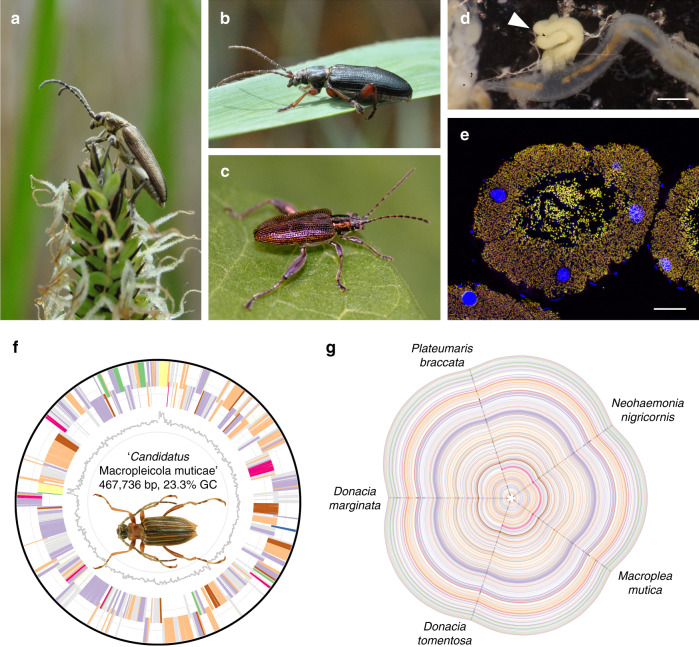
Symbiotic microbes can enable their host to access untapped nutritional resources but may also constrain niche space by promoting specialization. Here, we reconstruct functional changes in the evolutionary history of the symbiosis between a group of (semi-)aquatic herbivorous insects and mutualistic bacteria. Sequencing the symbiont genomes across 26 species of reed beetles (Chrysomelidae, Donaciinae) spanning four genera indicates that the genome-eroded mutualists provide life stage-specific benefits to larvae and adults, respectively. In the plant sap-feeding larvae, the symbionts are inferred to synthesize most of the essential amino acids as well as the B vitamin riboflavin. The adult reed beetles' folivory is likely supported by symbiont-encoded pectinases that complement the host-encoded set of cellulases, as revealed by transcriptome sequencing. However, mapping the occurrence of the symbionts' pectinase genes and the hosts' food plant preferences onto the beetles' phylogeny reveals multiple independent losses of pectinase genes in lineages that switched to feeding on pectin-poor plants, presumably constraining their hosts' subsequent adaptive potential.
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Douglas AE. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009;23:38–47.
-
- Feldhaar H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011;36:533–543.
-
- Sudakaran S, Kost C, Kaltenpoth M. Symbiont acquisition and replacement as a source of ecological innovation. Trends Microbiol. 2017;25:375–390. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
