

Metabolic Reprograming via Deletion of CISH in Human iPSC-Derived NK Cells Promotes In Vivo Persistence and Enhances Anti-tumor Activity
- PMID: 32531207
- PMCID: PMC7415618
- DOI: 10.1016/j.stem.2020.05.008
Metabolic Reprograming via Deletion of CISH in Human iPSC-Derived NK Cells Promotes In Vivo Persistence and Enhances Anti-tumor Activity
Abstract
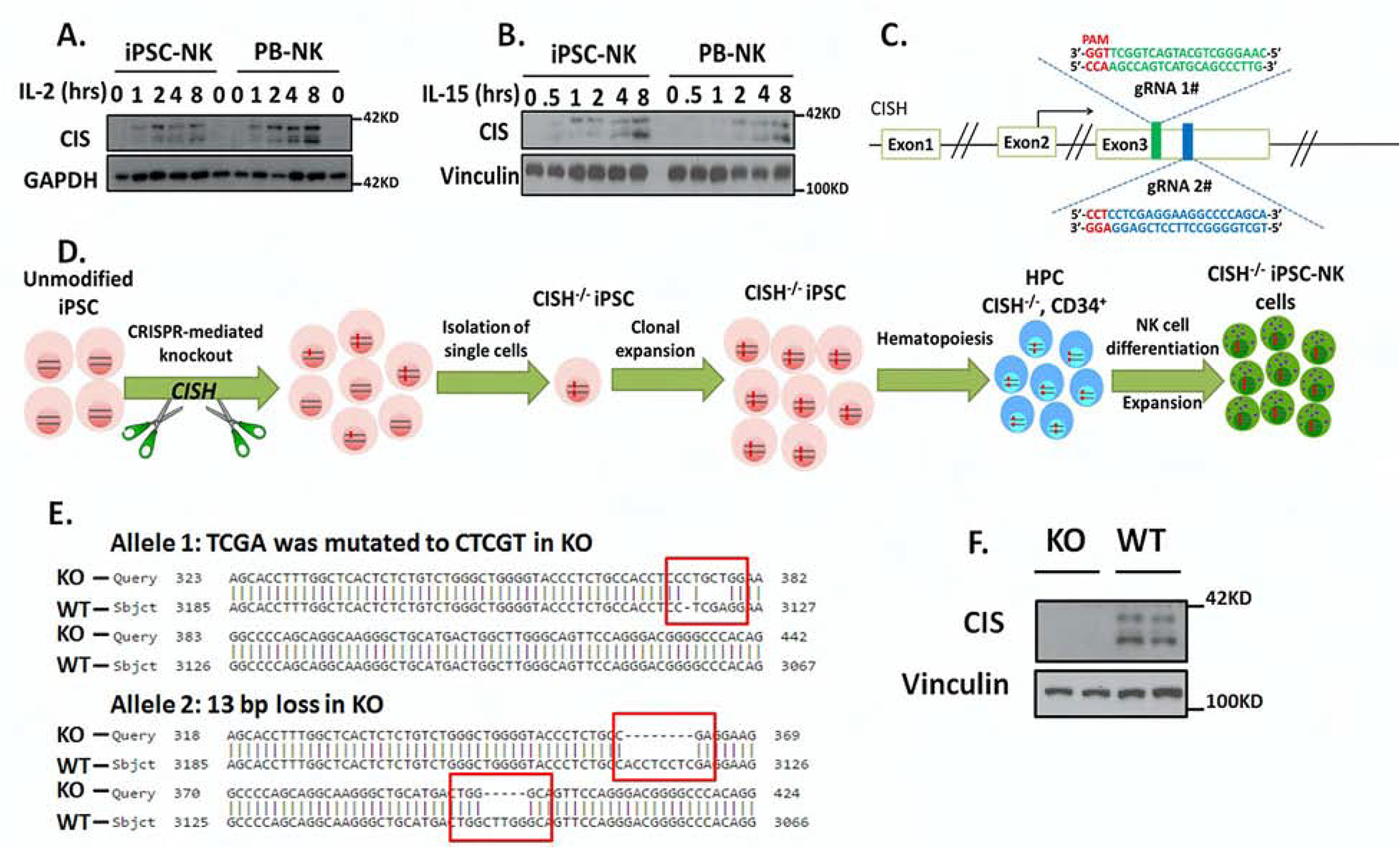
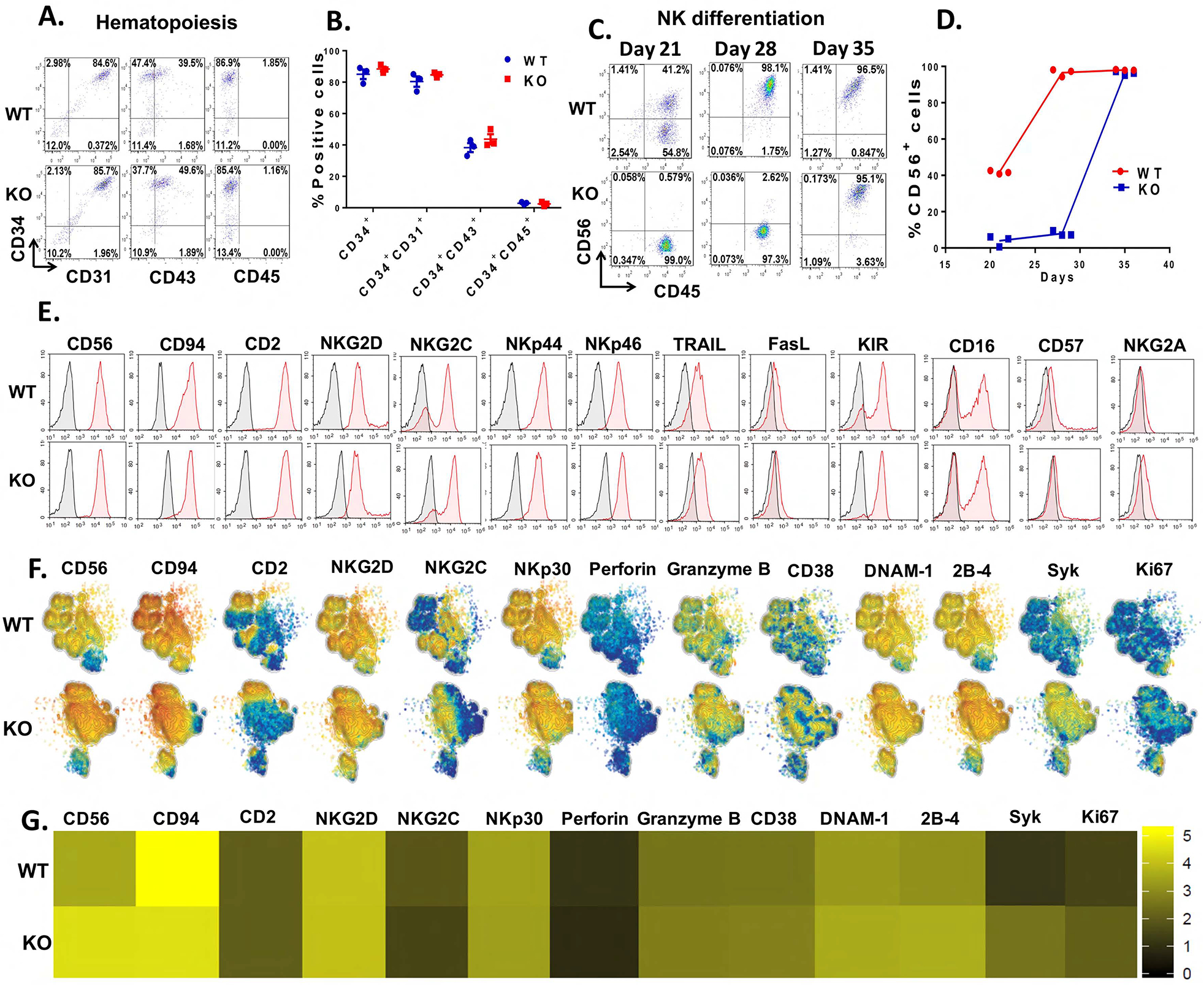
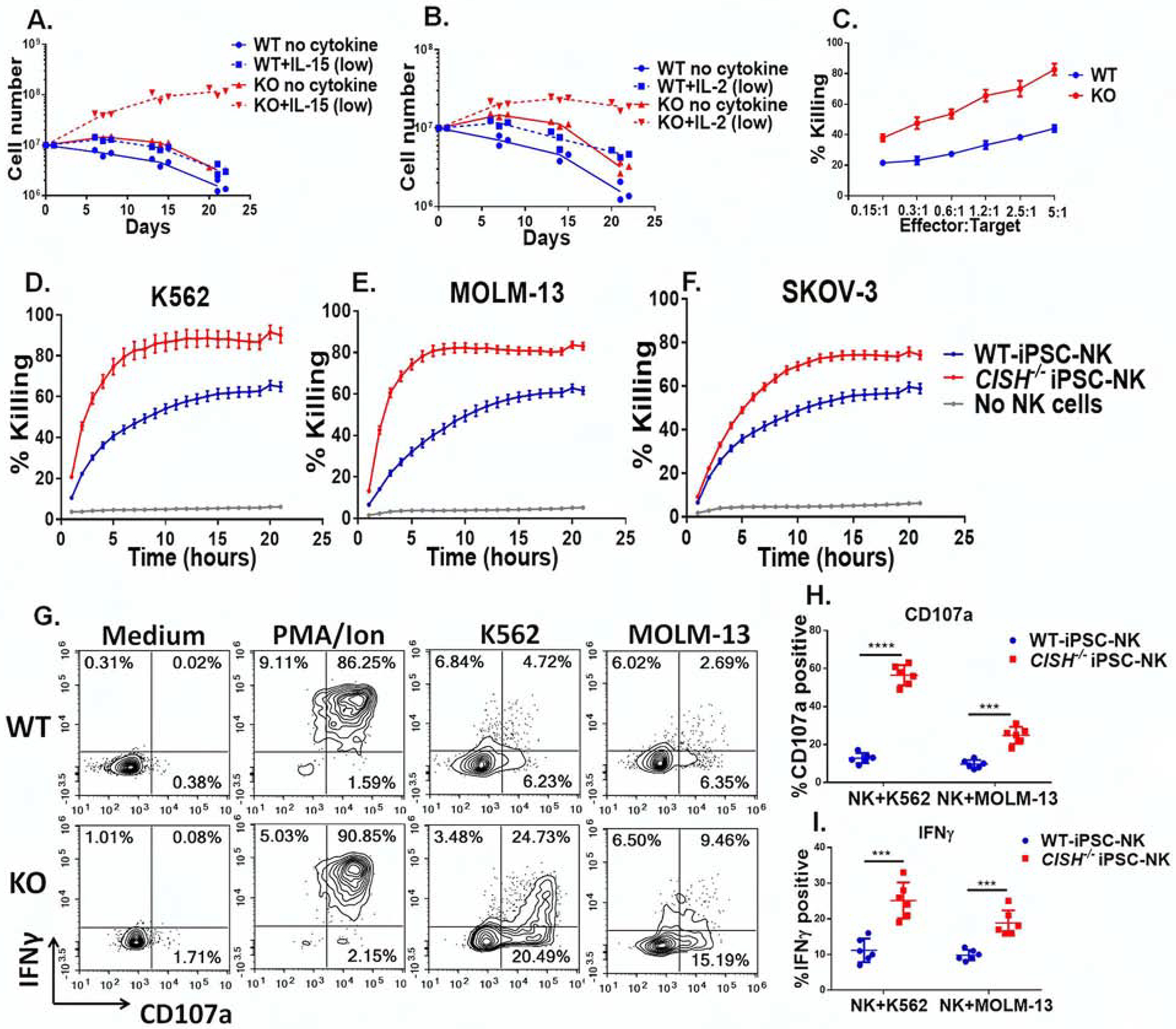
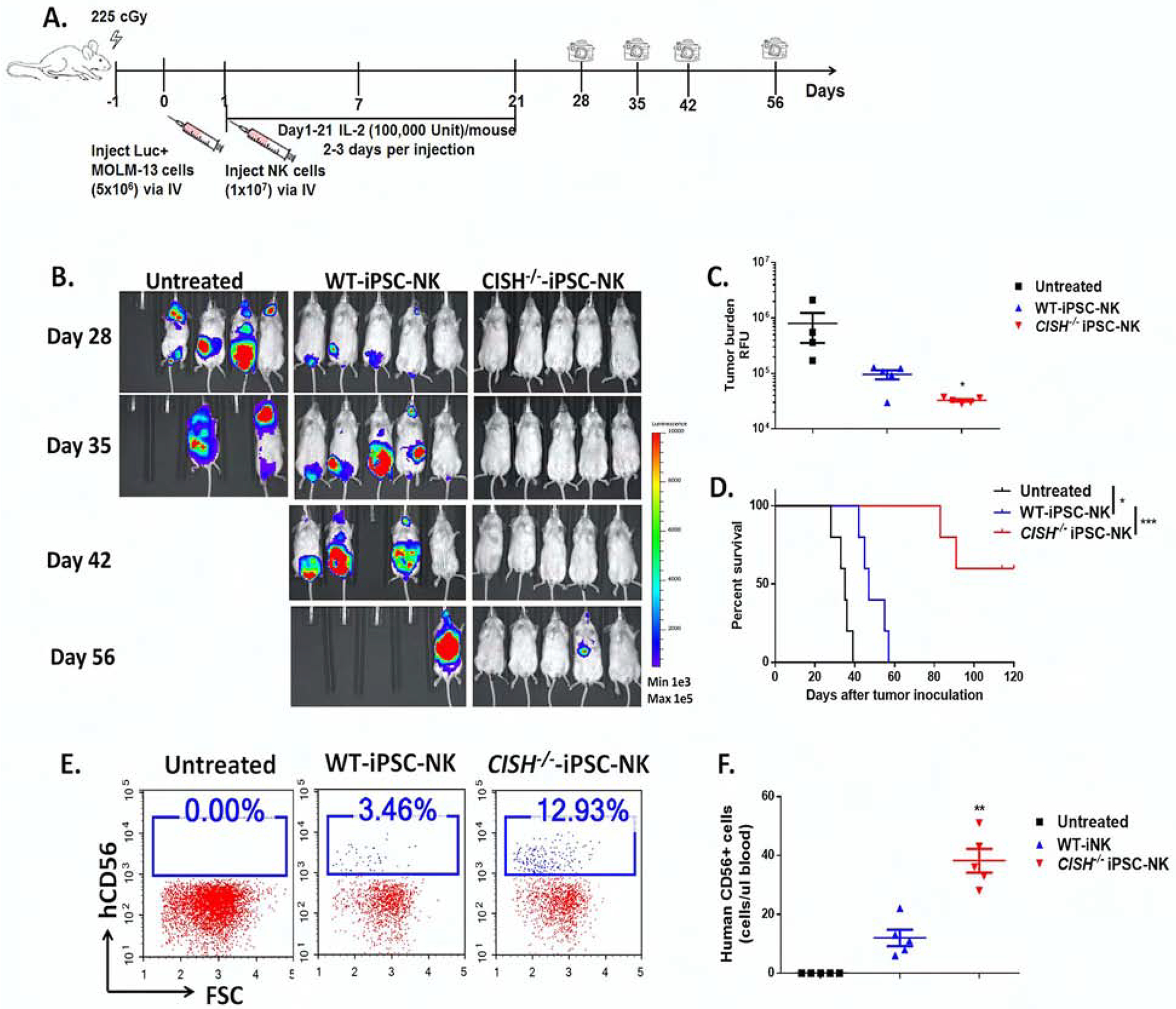
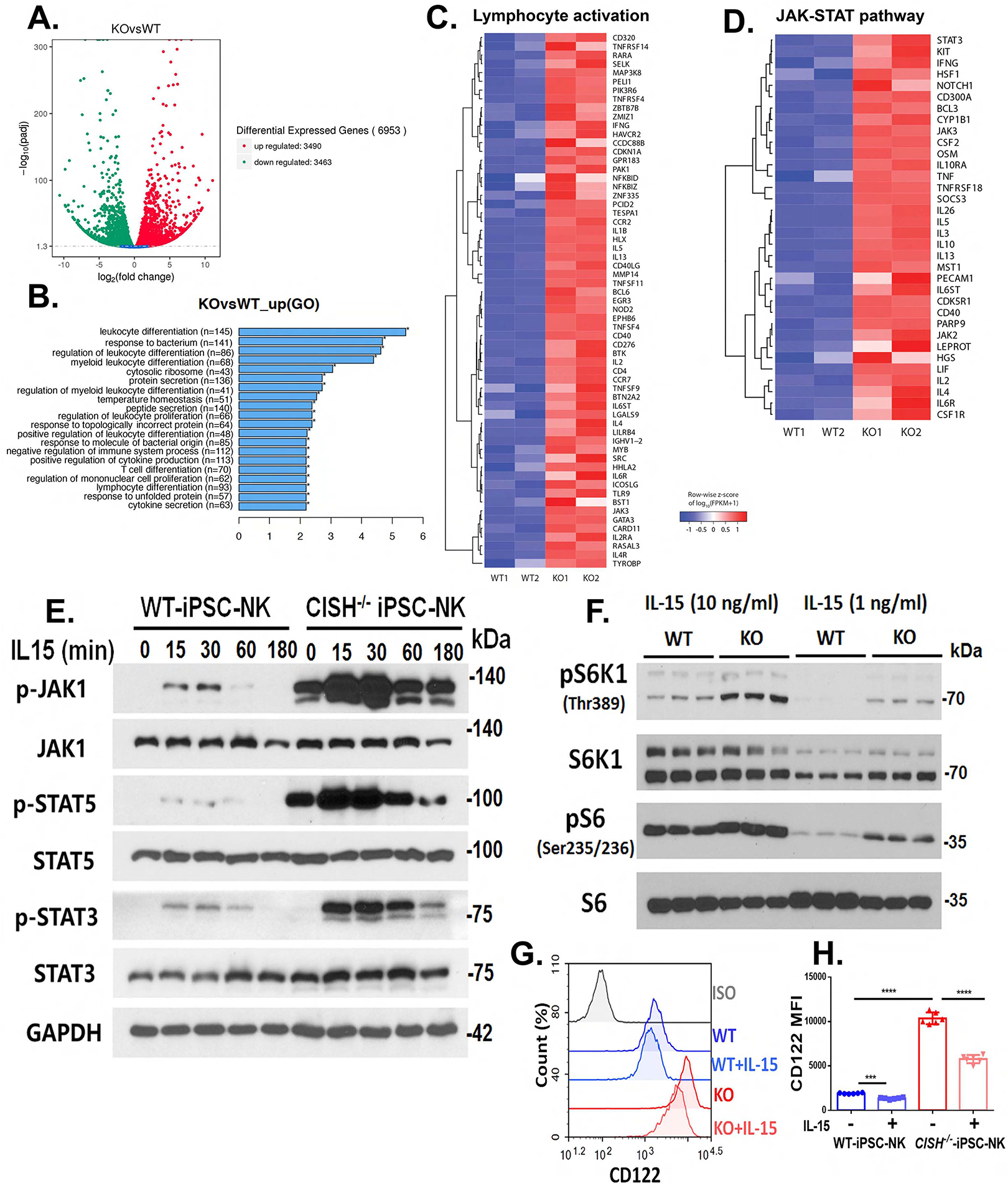
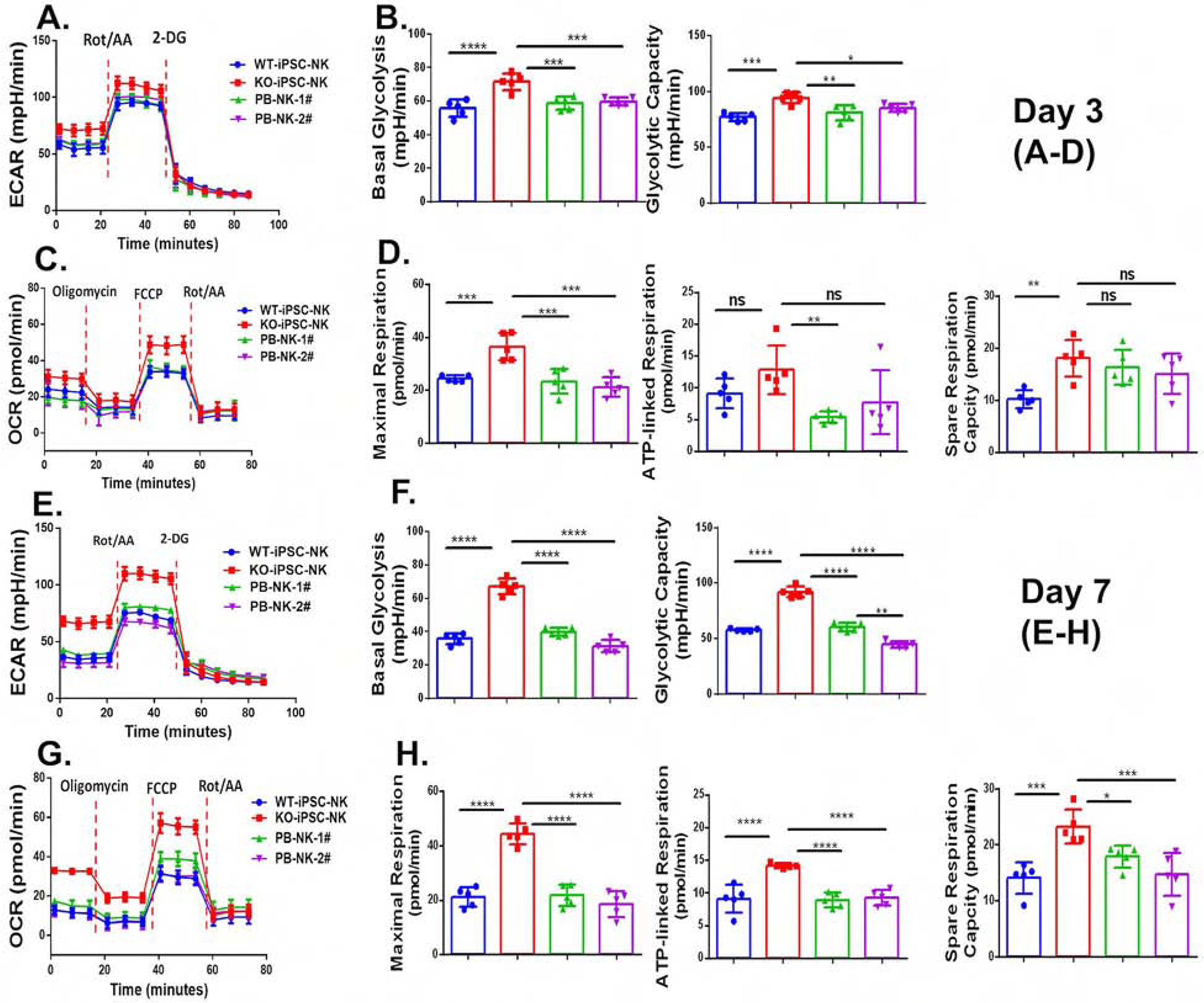
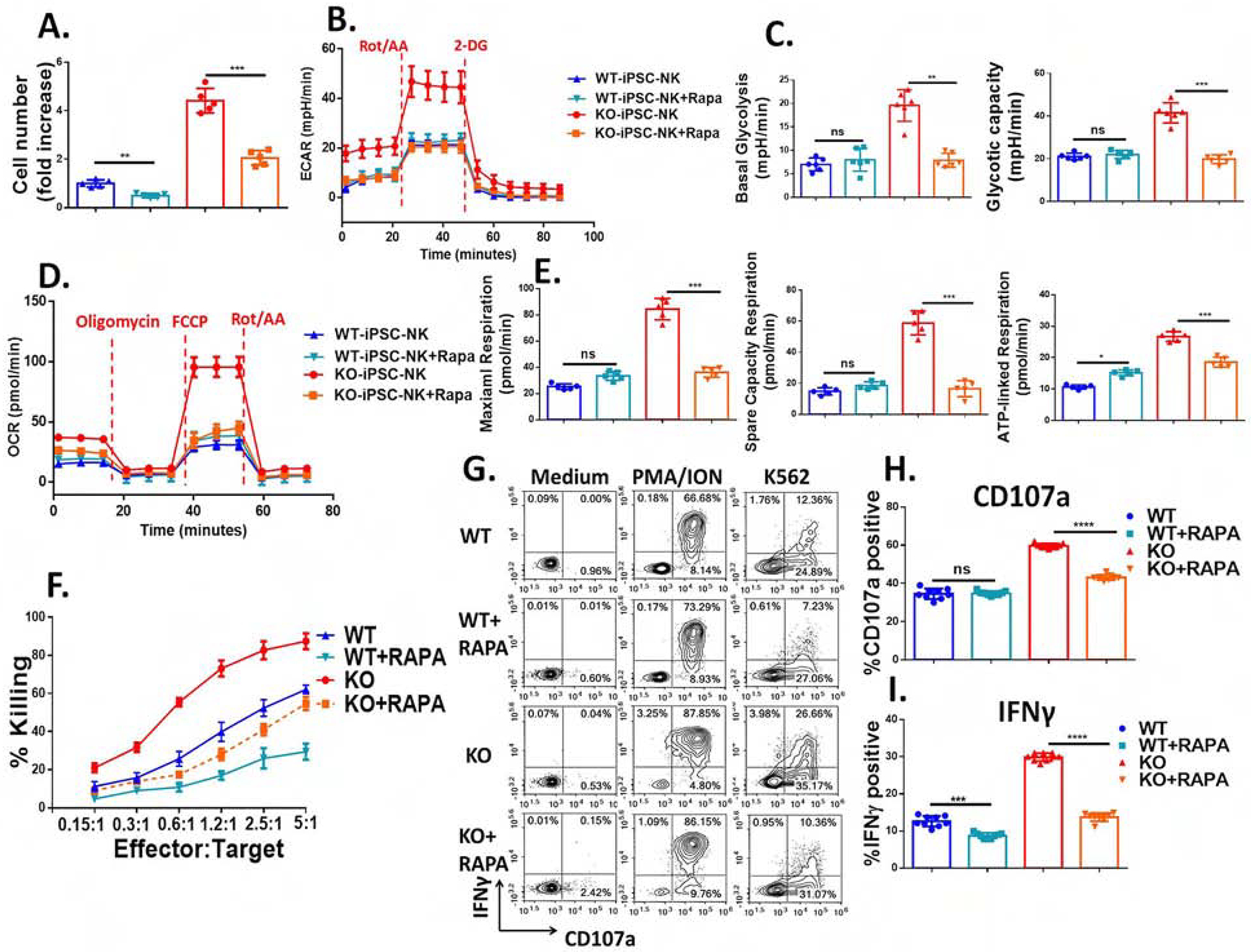
Cytokine-inducible SH2-containing protein (CIS; encoded by the gene CISH) is a key negative regulator of interleukin-15 (IL-15) signaling in natural killer (NK) cells. Here, we develop human CISH-knockout (CISH-/-) NK cells using an induced pluripotent stem cell-derived NK cell (iPSC-NK cell) platform. CISH-/- iPSC-NK cells demonstrate increased IL-15-mediated JAK-STAT signaling activity. Consequently, CISH-/- iPSC-NK cells exhibit improved expansion and increased cytotoxic activity against multiple tumor cell lines when maintained at low cytokine concentrations. CISH-/- iPSC-NK cells display significantly increased in vivo persistence and inhibition of tumor progression in a leukemia xenograft model. Mechanistically, CISH-/- iPSC-NK cells display improved metabolic fitness characterized by increased basal glycolysis, glycolytic capacity, maximal mitochondrial respiration, ATP-linked respiration, and spare respiration capacity mediated by mammalian target of rapamycin (mTOR) signaling that directly contributes to enhanced NK cell function. Together, these studies demonstrate that CIS plays a key role to regulate human NK cell metabolic activity and thereby modulate anti-tumor activity.
Keywords: CISH; IL-15; JAK-STAT; acute myelogenous leukemia; cell therapy; iPSCs; immunotherapy; mTOR; metabolic reprograming; natural killer cells.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests D.S.K. is a consultant for Fate Therapeutics, has equity, and receives income. The terms of this arrangement have been reviewed and approved by the University of California, San Diego in accordance with its conflict of interest policies. He also has patents filed or issued related to this work. K.-J.M. consults for and receives research support from Fate Therapeutics. K.-L.G. is co-founder and has an equity interest in Vivace Therapeutics, Inc., and OncoImmune, Inc. H.Z. has a patent filed related to this work.
Figures
References
-
- Assmann N, O’Brien KL, Donnelly RP, Dyck L, Zaiatz-Bittencourt V, Loftus RM, Heinrich P, Oefner PJ, Lynch L, Gardiner CM, et al. (2017). Srebp-controlled glucose metabolism is essential for NK cell functional responses. Nat Immunol 18, 1197–1206. - PubMed
-
- Cong JJ, Wang XW, Zheng XH, Wang D, Fu BQ, Sun R, Tian ZG, and Wei HM (2018). Dysfunction of Natural Killer Cells by FBP1-Induced Inhibition of Glycolysis during Lung Cancer Progression. Cell Metab 28, 243–255.e245. - PubMed
-
- Delconte RB, Kolesnik TB, Dagley LF, Rautela J, Shi W, Putz EM, Stannard K, Zhang JG, Teh C, Firth M, et al. (2016). CIS is a potent checkpoint in NK cell-mediated tumor immunity. Nat Immunol 17, 816–824. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
