

Gene-environment regulation of chamber-specific maturation during hypoxemic perinatal circulatory transition
- PMID: 32533200
- PMCID: PMC7843236
- DOI: 10.1007/s00109-020-01933-8
Gene-environment regulation of chamber-specific maturation during hypoxemic perinatal circulatory transition
Abstract
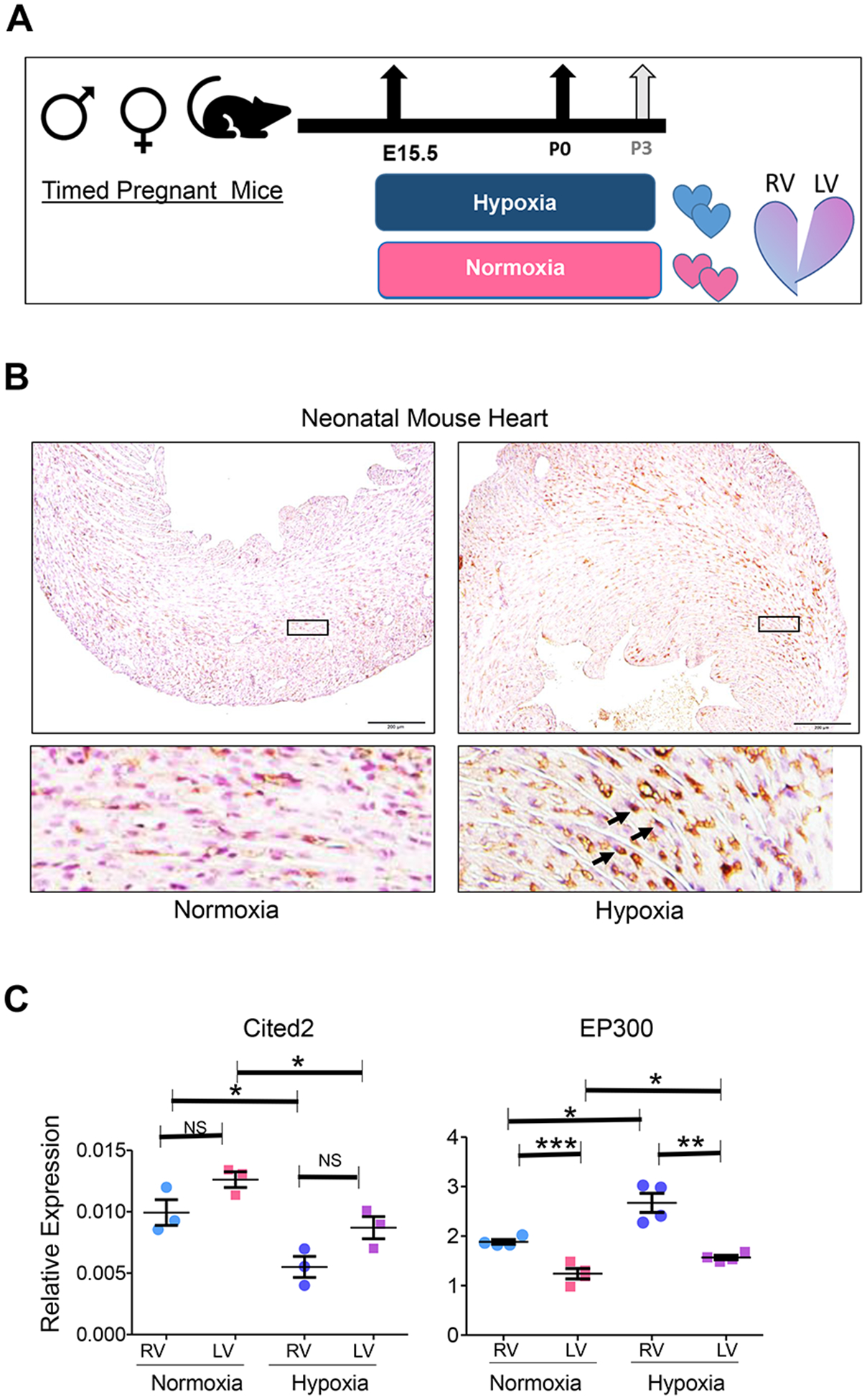
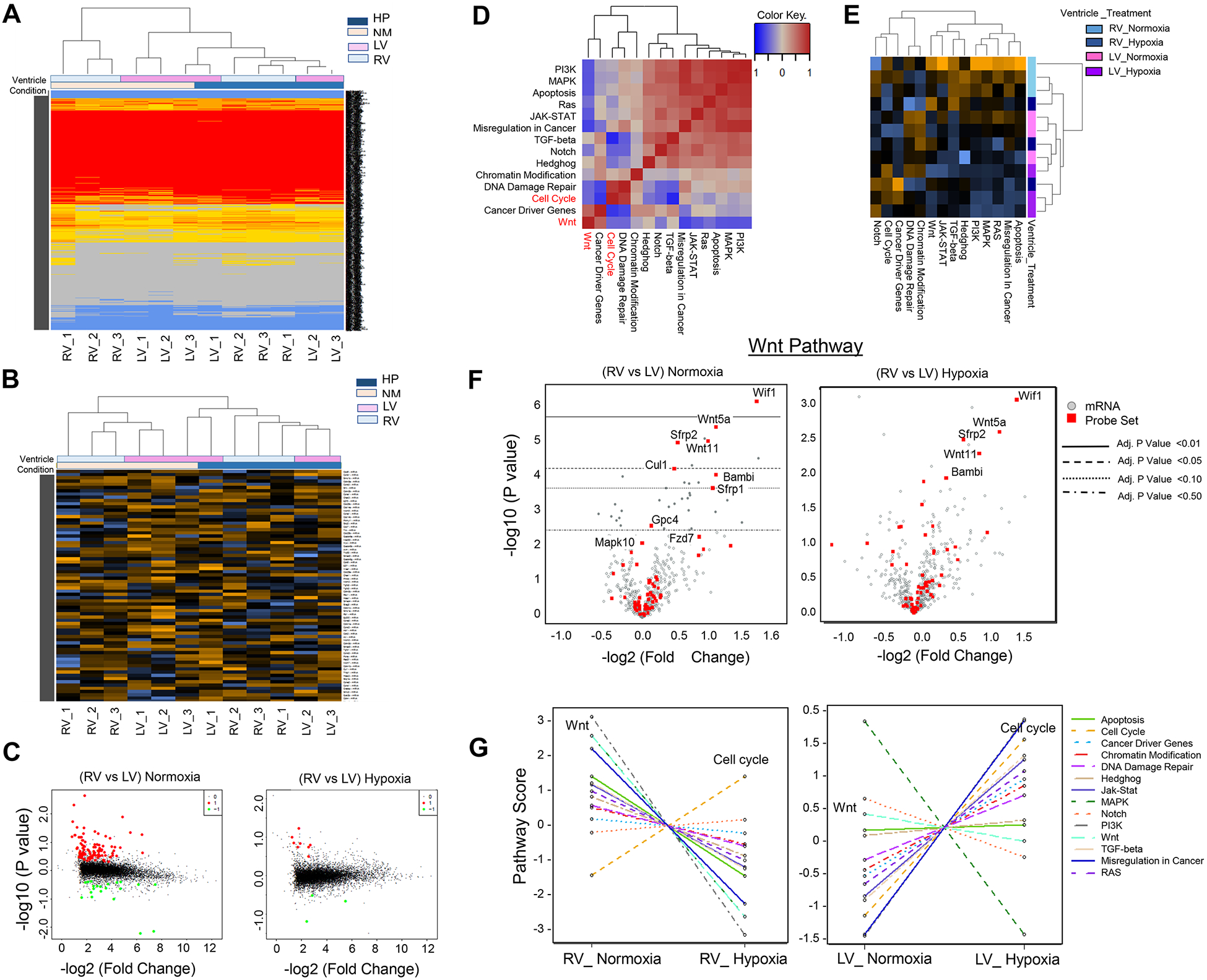
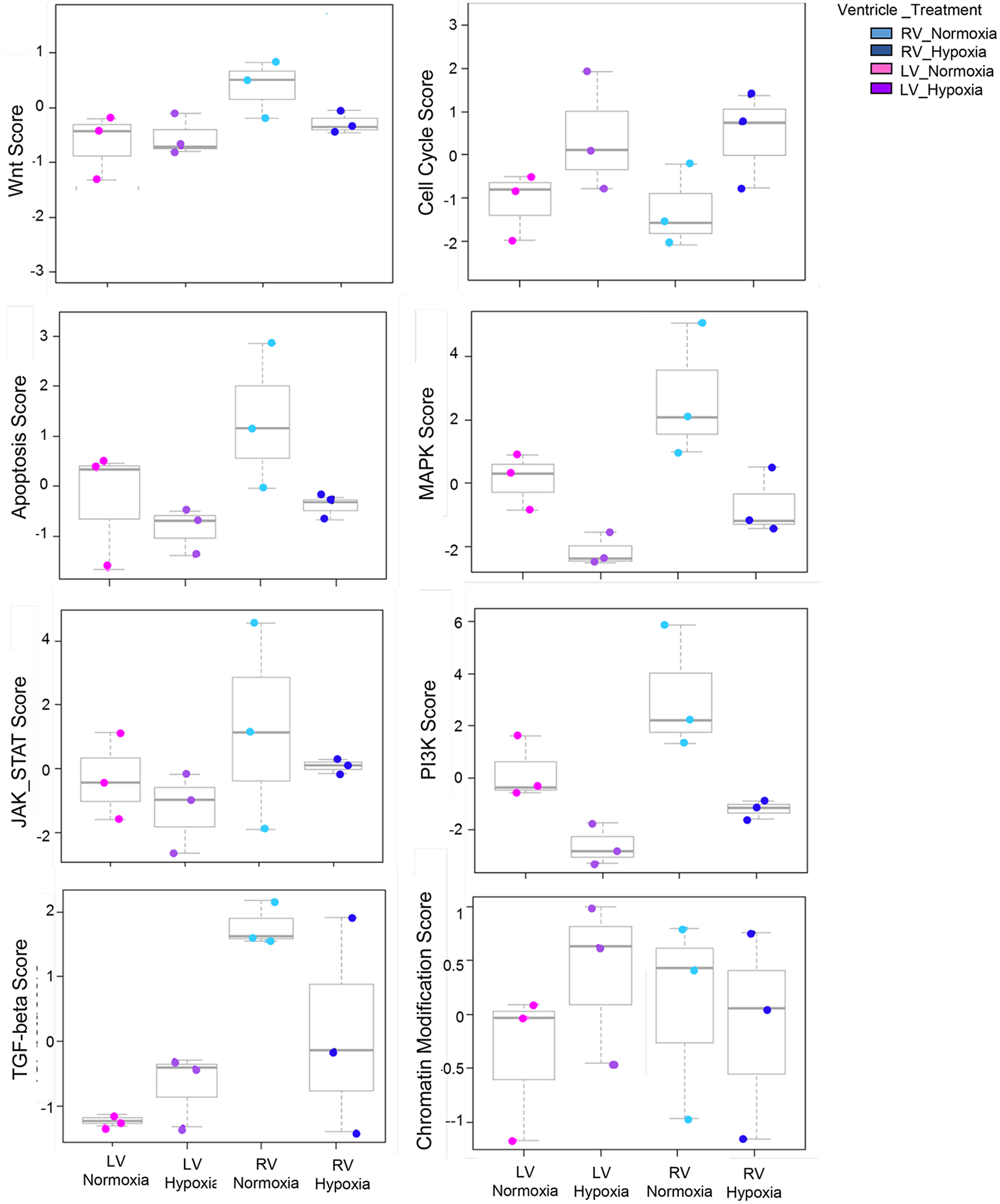
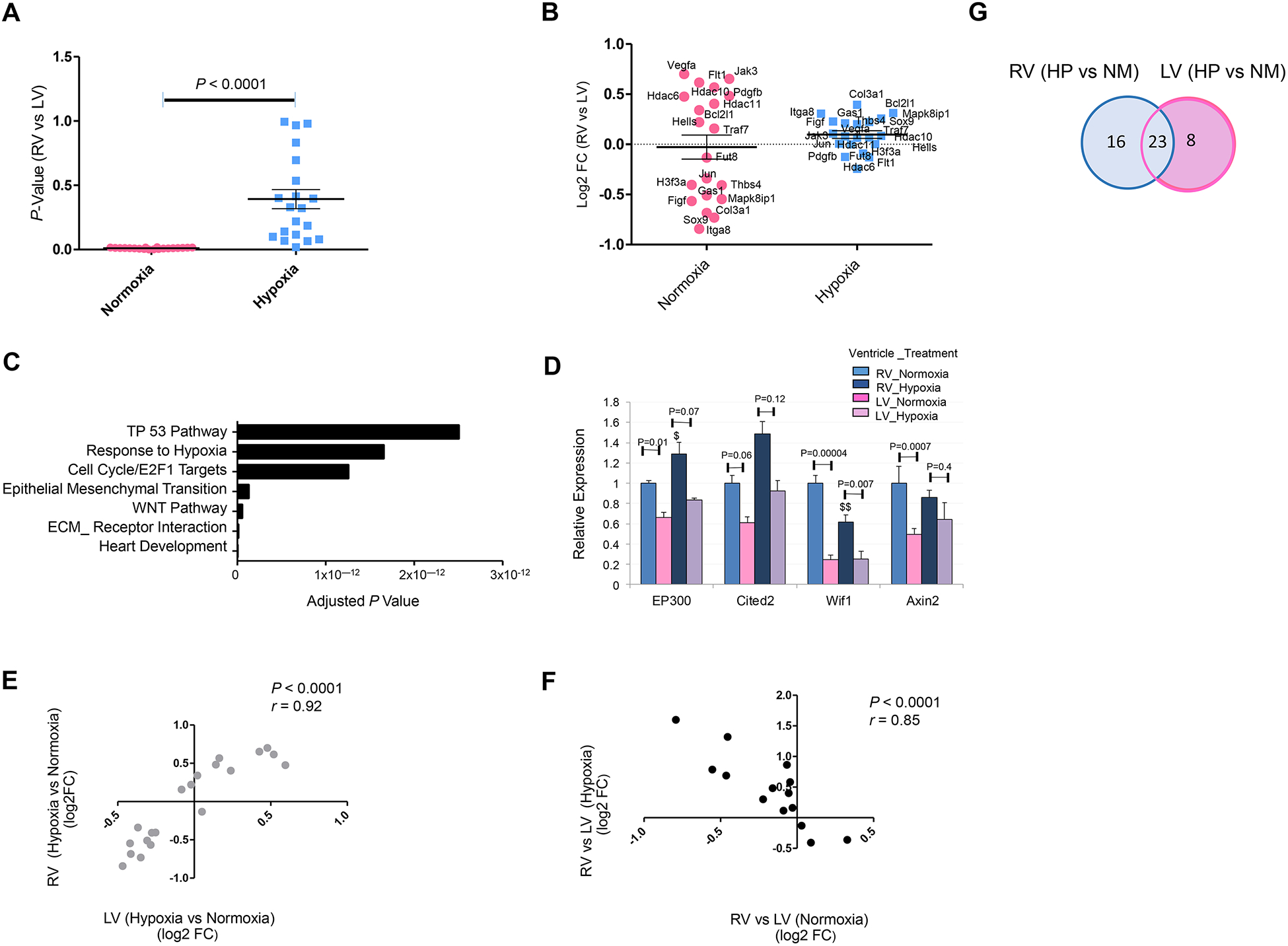
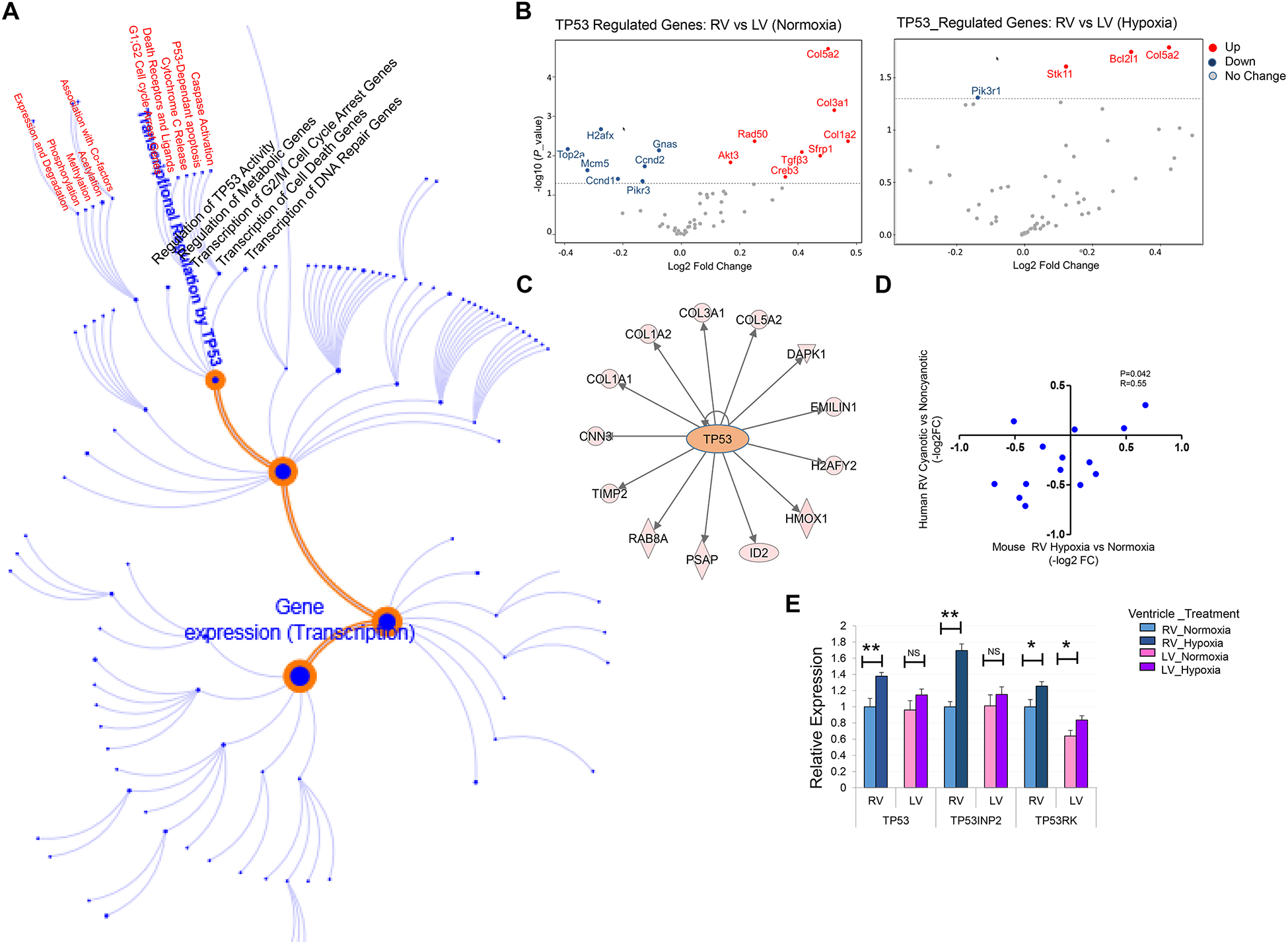
Chamber-specific and temporally regulated perinatal cardiac growth and maturation is critical for functional adaptation of the heart and may be altered significantly in response to perinatal stress, such as systemic hypoxia (hypoxemia), leading to significant pathology, even mortality. Understanding transcriptome regulation of neonatal heart chambers in response to hypoxemia is necessary to develop chamber-specific therapies for infants with cyanotic congenital heart defects (CHDs). We sought to determine chamber-specific transcriptome programming during hypoxemic perinatal circulatory transition. We performed transcriptome-wide analysis on right ventricle (RV) and left ventricle (LV) of postnatal day 3 (P3) mouse hearts exposed to perinatal hypoxemia. Hypoxemia decreased baseline differences between RV and LV leading to significant attenuation of ventricular patterning (AVP), which involved several molecular pathways, including Wnt signaling suppression and cell cycle induction. Notably, robust changes in RV transcriptome in hypoxemic condition contributed significantly to the AVP. Remarkably, suppression of epithelial mesenchymal transition (EMT) and dysregulation of the TP53 signaling were prominent hallmarks of the AVP genes in neonatal mouse heart. Furthermore, members of the TP53-related gene family were dysregulated in the hypoxemic RVs of neonatal mouse and cyanotic Tetralogy of Fallot hearts. Integrated analysis of chamber-specific transcriptome revealed hypoxemia-specific changes that were more robust in RVs compared with LVs, leading to previously uncharacterized AVP induced by perinatal hypoxemia. Remarkably, reprogramming of EMT process and dysregulation of the TP53 network contributed to transcriptome remodeling of neonatal heart during hypoxemic circulatory transition. These insights may enhance our understanding of hypoxemia-induced pathogenesis in newborn infants with cyanotic CHD phenotypes. KEY MESSAGES: During perinatal circulatory transition, transcriptome programming is a major driving force of cardiac chamber-specific maturation and adaptation to hemodynamic load and external environment. During hypoxemic perinatal transition, transcriptome reprogramming may affect chamber-specific growth and development, particularly in newborns with congenital heart defects (CHDs). Chamber-specific transcriptome changes during hypoxemic perinatal transition are yet to be fully elucidated. Systems-based analysis of hypoxemic neonatal hearts at postnatal day 3 reveals chamber-specific transcriptome signatures during hypoxemic perinatal transition, which involve attenuation of ventricular patterning (AVP) and repression of epithelial mesenchymal transition (EMT). Key regulatory circuits involved in hypoxemia response were identified including suppression of Wnt signaling, induction of cellular proliferation and dysregulation of TP53 network.
Keywords: Congenital heart defects; Hypoxia; Neonatal heart maturation; Tetralogy of Fallot; Transcriptome.
Conflict of interest statement
CONFLICT OF INTEREST:
None.
Figures
References
-
- Zhao Y, Kang X, Gao F, Guzman A, Lau RP, Biniwale R, Wadehra M, Reemtsen B, Garg M, Halnon N, Quintero-Rivera F, Arsdell GV, Giovanni C, Nelson SF, Touma M & the UCLA Congenital Heart Defects BioCore Faculty. Gene-Environment Regulatory Circuits of Right Ventricular Pathology in Tetralogy of Fallot JMolMed Ms. No. JMME-D-19–00563R1. In press. - PMC - PubMed
-
- Finnemore A, Groves A. Physiology of the fetal and transitional circulation. Semin Fetal Neonatal Med. 2015;20(4):210–216. - PubMed
-
- Rudolph AM. Myocardial growth before and after birth: clinical implications. Acta Paediatr. 2000; 89(2):129–133. - PubMed
-
- Sinha SK, Donn SM. Fetal-to-neonatal maladaptation. Semin Fetal Neonatal Med. 2006;11(3):166–173. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
