

Auditory representation of learned sound sequences in motor regions of the macaque brain
- PMID: 32541016
- PMCID: PMC7334521
- DOI: 10.1073/pnas.1915610117
Auditory representation of learned sound sequences in motor regions of the macaque brain
Abstract
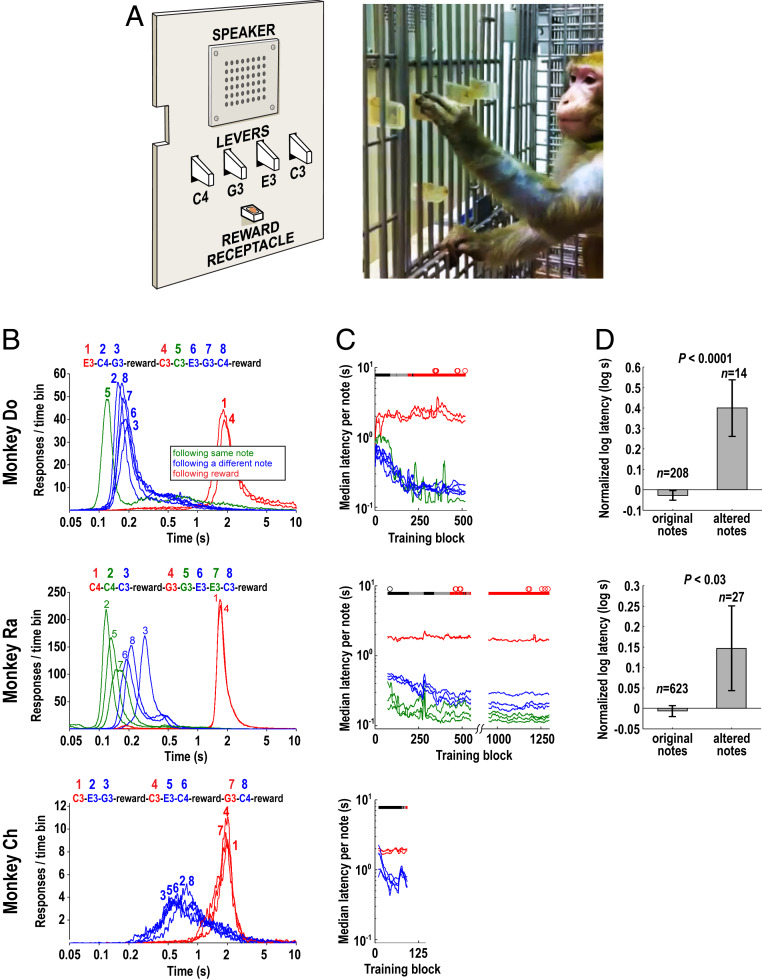
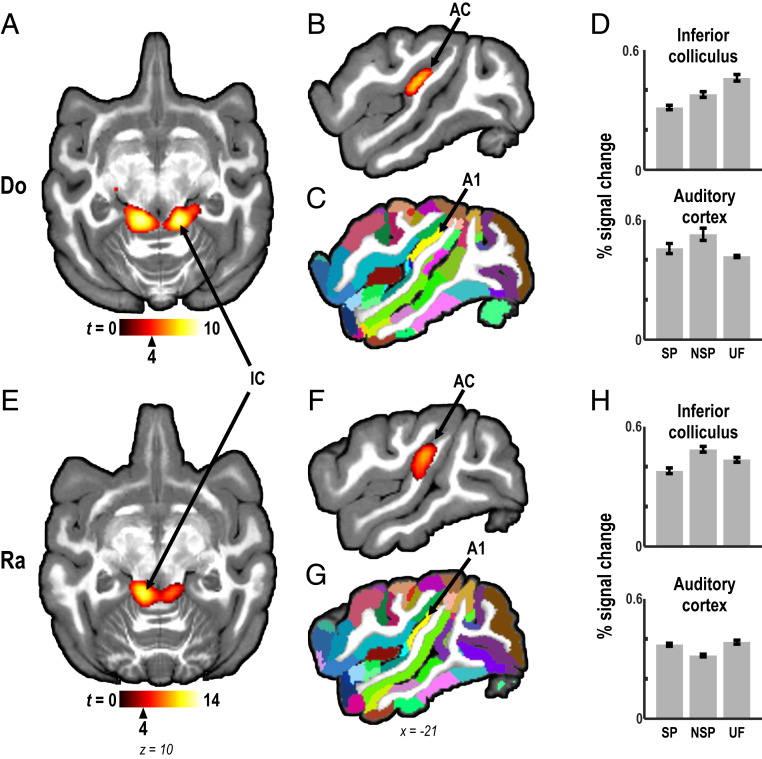
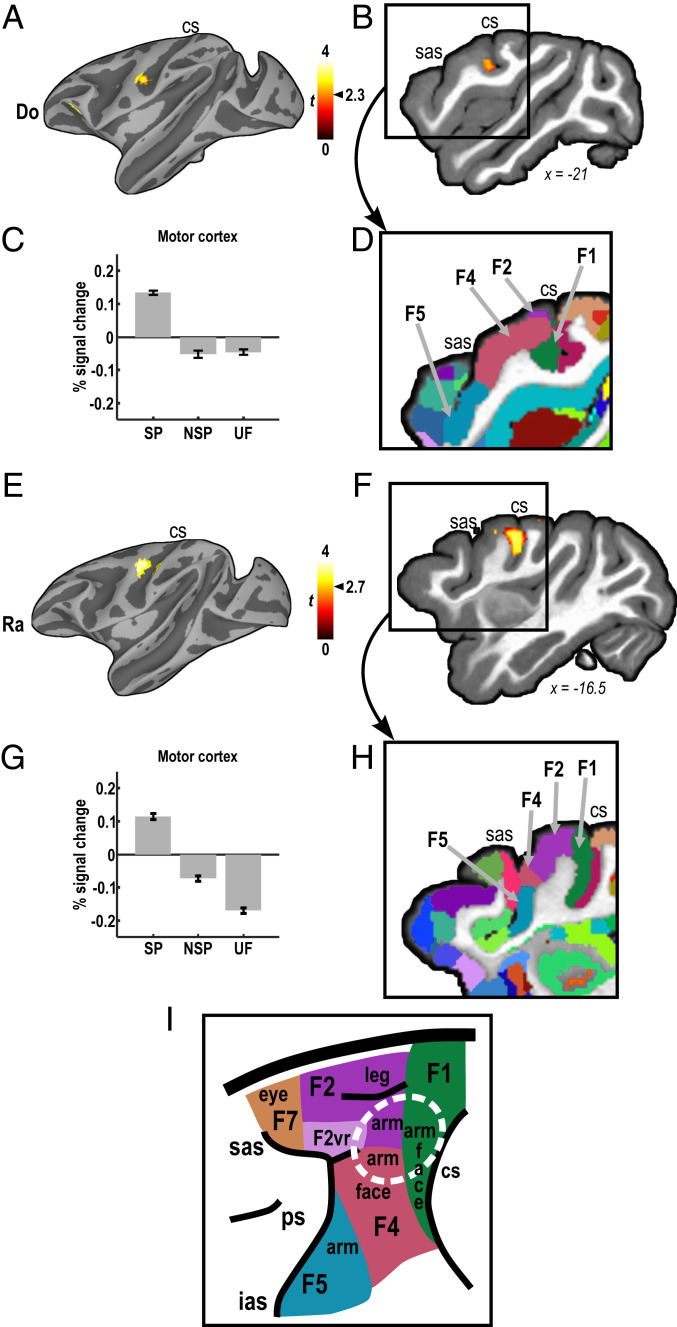
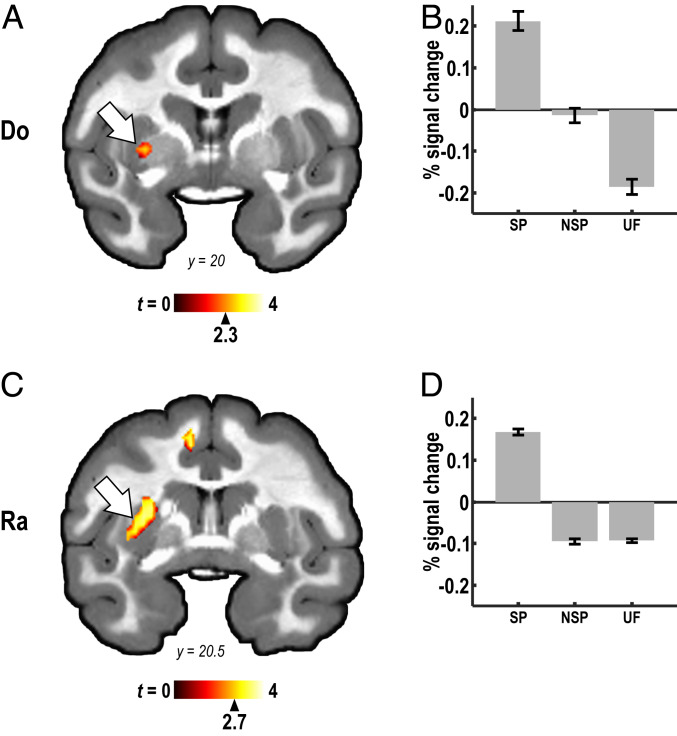
Human speech production requires the ability to couple motor actions with their auditory consequences. Nonhuman primates might not have speech because they lack this ability. To address this question, we trained macaques to perform an auditory-motor task producing sound sequences via hand presses on a newly designed device ("monkey piano"). Catch trials were interspersed to ascertain the monkeys were listening to the sounds they produced. Functional MRI was then used to map brain activity while the animals listened attentively to the sound sequences they had learned to produce and to two control sequences, which were either completely unfamiliar or familiar through passive exposure only. All sounds activated auditory midbrain and cortex, but listening to the sequences that were learned by self-production additionally activated the putamen and the hand and arm regions of motor cortex. These results indicate that, in principle, monkeys are capable of forming internal models linking sound perception and production in motor regions of the brain, so this ability is not special to speech in humans. However, the coupling of sounds and actions in nonhuman primates (and the availability of an internal model supporting it) seems not to extend to the upper vocal tract, that is, the supralaryngeal articulators, which are key for the production of speech sounds in humans. The origin of speech may have required the evolution of a "command apparatus" similar to the control of the hand, which was crucial for the evolution of tool use.
Keywords: auditory cortex; internal models; motor cortex; putamen.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Levelt W. J. M., Speaking: From Intention to Articulation (MIT Press, 1989).
-
- Cusack R., Wild C. J., Zubiaurre-Elorza L., Linke A. C., Why does language not emerge until the second year? Hear. Res. 366, 75–81 (2018). - PubMed
-
- Jordan M. I., Rumelhart D. E., Forward models: Supervised learning with a distal teacher. Cogn. Sci. 16, 307–354 (1992).
-
- Smith C. R., Residual hearing and speech production in deaf children. J. Speech Hear. Res. 18, 795–811 (1975). - PubMed
-
- von Holst E., Mittelstaedt H., Das Reafferenzprinzip–Wechselwirkungen zwischen Zentralnervensystem und Peripherie. Naturwissenschaften 37, 464–476 (1950).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
