

Clonally expanding smooth muscle cells promote atherosclerosis by escaping efferocytosis and activating the complement cascade
- PMID: 32541024
- PMCID: PMC7354942
- DOI: 10.1073/pnas.2006348117
Clonally expanding smooth muscle cells promote atherosclerosis by escaping efferocytosis and activating the complement cascade
Abstract
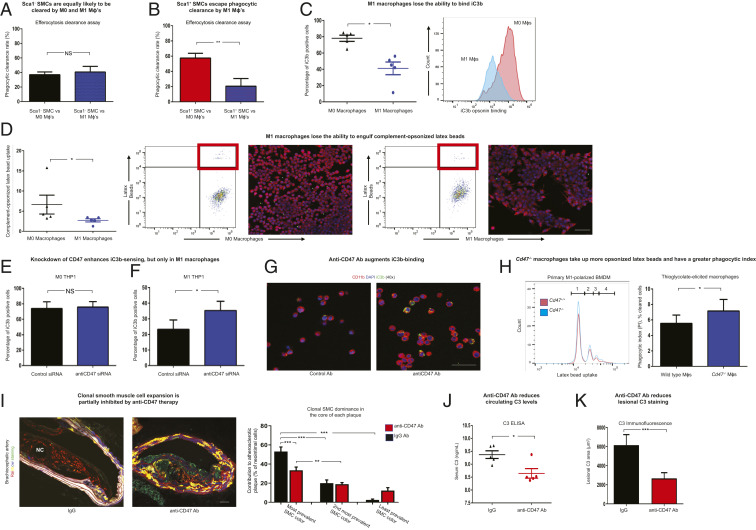
Atherosclerosis is the process underlying heart attack and stroke. Despite decades of research, its pathogenesis remains unclear. Dogma suggests that atherosclerotic plaques expand primarily via the accumulation of cholesterol and inflammatory cells. However, recent evidence suggests that a substantial portion of the plaque may arise from a subset of "dedifferentiated" vascular smooth muscle cells (SMCs) which proliferate in a clonal fashion. Herein we use multicolor lineage-tracing models to confirm that the mature SMC can give rise to a hyperproliferative cell which appears to promote inflammation via elaboration of complement-dependent anaphylatoxins. Despite being extensively opsonized with prophagocytic complement fragments, we find that this cell also escapes immune surveillance by neighboring macrophages, thereby exacerbating its relative survival advantage. Mechanistic studies indicate this phenomenon results from a generalized opsonin-sensing defect acquired by macrophages during polarization. This defect coincides with the noncanonical up-regulation of so-called don't eat me molecules on inflamed phagocytes, which reduces their capacity for programmed cell removal (PrCR). Knockdown or knockout of the key antiphagocytic molecule CD47 restores the ability of macrophages to sense and clear opsonized targets in vitro, allowing for potent and targeted suppression of clonal SMC expansion in the plaque in vivo. Because integrated clinical and genomic analyses indicate that similar pathways are active in humans with cardiovascular disease, these studies suggest that the clonally expanding SMC may represent a translational target for treating atherosclerosis.
Keywords: CD47; atherosclerosis; clonality; efferocytosis; smooth muscle cells.
Conflict of interest statement
Competing interest statement: I.L.W. and N.J.L. are cofounders of Forty Seven, Inc., an immunooncology company. This company was recently acquired by Gilead Sciences; the purchase did not include stock in Gilead. I.L.W. and N.J.L. do not currently have any consulting agreement with Gilead Sciences.
Figures



Similar articles
-
Smooth muscle cell-specific CD47 deletion suppresses atherosclerosis.Life Sci. 2025 Jan 15;361:123315. doi: 10.1016/j.lfs.2024.123315. Epub 2024 Dec 13. Life Sci. 2025. PMID: 39675550
-
EphA2 Expression Regulates Inflammation and Fibroproliferative Remodeling in Atherosclerosis.Circulation. 2017 Aug 8;136(6):566-582. doi: 10.1161/CIRCULATIONAHA.116.026644. Epub 2017 May 9. Circulation. 2017. PMID: 28487392 Free PMC article.
-
2021 Jeffrey M. Hoeg Award Lecture: Defining the Role of Efferocytosis in Cardiovascular Disease: A Focus on the CD47 (Cluster of Differentiation 47) Axis.Arterioscler Thromb Vasc Biol. 2022 Jun;42(6):e145-e154. doi: 10.1161/ATVBAHA.122.317049. Epub 2022 Apr 7. Arterioscler Thromb Vasc Biol. 2022. PMID: 35387480 Free PMC article. Review.
-
Efferocytosis in atherosclerotic lesions: Malfunctioning regulatory pathways and control mechanisms.Pharmacol Ther. 2018 Aug;188:12-25. doi: 10.1016/j.pharmthera.2018.02.003. Epub 2018 Feb 11. Pharmacol Ther. 2018. PMID: 29444453 Review.
-
Mechanism of efferocytosis in atherosclerosis.J Mol Med (Berl). 2024 Jul;102(7):831-840. doi: 10.1007/s00109-024-02439-3. Epub 2024 May 10. J Mol Med (Berl). 2024. PMID: 38727748 Review.
Cited by
-
Immunotherapy-Associated Atherosclerosis: A Comprehensive Review of Recent Findings and Implications for Future Research.Curr Treat Options Cardiovasc Med. 2023;25(12):715-735. doi: 10.1007/s11936-023-01024-0. Epub 2023 Dec 15. Curr Treat Options Cardiovasc Med. 2023. PMID: 38213548 Free PMC article. Review.
-
The immune system in cardiovascular diseases: from basic mechanisms to therapeutic implications.Signal Transduct Target Ther. 2025 May 23;10(1):166. doi: 10.1038/s41392-025-02220-z. Signal Transduct Target Ther. 2025. PMID: 40404619 Free PMC article. Review.
-
Mechanisms of vascular smooth muscle cell investment and phenotypic diversification in vascular diseases.Biochem Soc Trans. 2021 Nov 1;49(5):2101-2111. doi: 10.1042/BST20210138. Biochem Soc Trans. 2021. PMID: 34495326 Free PMC article. Review.
-
Clinical implications of inflammation in atheroma formation and novel therapies in cardiovascular diseases.Front Cell Dev Biol. 2023 Mar 16;11:1148768. doi: 10.3389/fcell.2023.1148768. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37009489 Free PMC article. Review.
-
Wnt16 Promotes Vascular Smooth Muscle Contractile Phenotype and Function via Taz (Wwtr1) Activation in Male LDLR-/- Mice.Endocrinology. 2023 Dec 23;165(2):bqad192. doi: 10.1210/endocr/bqad192. Endocrinology. 2023. PMID: 38123514 Free PMC article.
References
-
- World Health Organization , The top 10 causes of death (2018). https://www.who.int/en/news-room/fact-sheets/detail/the-top-10-causes-of.... Accessed 20 January 2020.
-
- Libby P., Ridker P. M., Hansson G. K., Progress and challenges in translating the biology of atherosclerosis. Nature 473, 317–325 (2011). - PubMed
-
- Ross R., Atherosclerosis—An inflammatory disease. N. Engl. J. Med. 340, 115–126 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
