

Review
doi: 10.1002/alz.12084.
Epub 2020 Jun 16.
The biogenesis and biology of amyloid β oligomers in the brain
Affiliations
- PMID: 32543725
- PMCID: PMC7984270
- DOI: 10.1002/alz.12084
Item in Clipboard
Review
The biogenesis and biology of amyloid β oligomers in the brain
Alzheimers Dement.
2020 Nov.
Erratum in
-
Erratum.Alzheimers Dement. 2023 Jan 2. doi: 10.1002/alz.12791. Online ahead of print. Alzheimers Dement. 2023. PMID: 36591792 No abstract available.
Abstract
The repeated failure of clinical trials targeting the amyloid beta (Aβ) protein has challenged the amyloid cascade hypothesis. In this perspective, I discuss the biogenesis and biology of Aβ, from the arrangement of its atoms to its effects on the human brain. I hope that this analysis will help guide future attempts to home in on this elusive therapeutic target.
© 2020 The Authors. Alzheimer's & Dementia published by Wiley Periodicals LLC on behalf of Alzheimer's Association.
Conflict of interest statement
No conflicts of interest
Figures
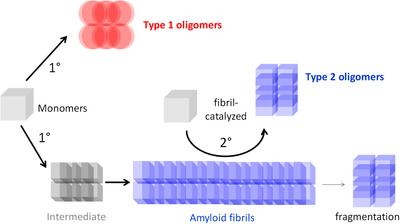
Primary and secondary nucleation of Aβ oligomers. Adapted, with permission, from Liu et al., “Quaternary structure defines a large class of Aβ oligomers neutralized by sequestration” Cell Reports, 2015. Type 1 oligomers arise via primary nucleation. Type 2 oligomers arise via fibril‐catalyzed secondary nucleation. Both types of oligomers compete for a common pool of monomers
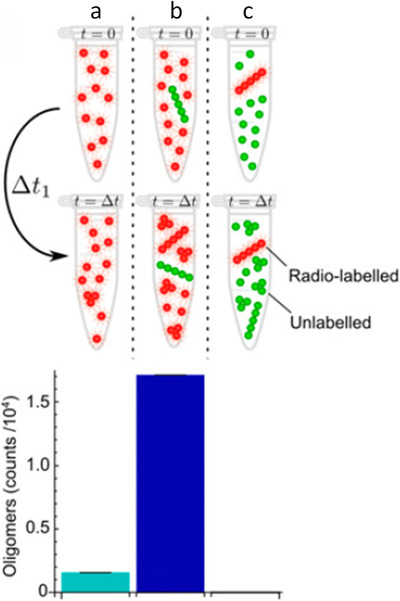
Fibril‐catalyzed secondary nucleation is the favored reaction process. Adapted, with permission, from Cohen et al., “Proliferation of amyloid‐β42 aggregates occurs through a secondary nucleation mechanism” PNAS, 2013. Radio‐labeled monomers (reaction a), radiolabelled monomers plus unlabeled fibrils (reaction b), or unlabeled monomers plus radio‐labeled fibrils (reaction c) were incubated for a period of time, Δt. The resultant oligomers in the reaction mixture were isolated by size‐exclusion chromatography, and the amount of radioactivity in the oligomer fractions was measured. The quantity of oligomers generated by secondary nucleation (reaction b) greatly exceeds that generated by primary nucleation (reaction a) or by fragmentation (reaction c)
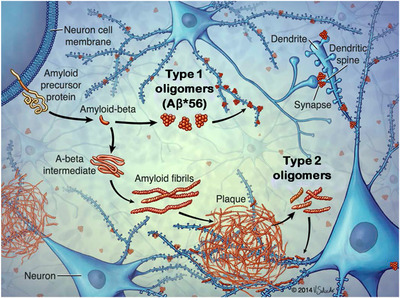
Biogenesis of types 1 and 2 Aβ oligomers in the brain. Adapted, with permission, from Liu et al., “Quaternary structure defines a large class of Aβ oligomers neutralized by sequestration” Cell Reports, 2015. Type 1 oligomers form by primary nucleation, do not have basic the structural feature of amyloid fibrils (ie, do not contain in‐register β‐sheets), are not concentrated around amyloid plaque cores, and appear before amyloid plaques.
24
Aβ*56
10
is a type 1 oligomer.
24
Type 2 oligomers form by secondary nucleation, contain in‐register β‐sheet structure, are concentrated around amyloid plaque cores, and appear after amyloid plaques.
24
Both types of oligomers affect neuronal signaling pathways through interactions with dendritic spines
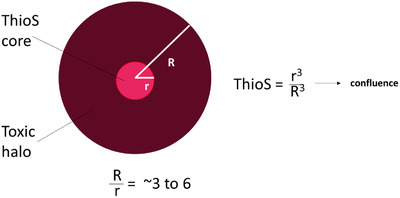
Type 2 oligomers reach confluence when the Thioflavin S plaque load is ≈ 0.5% to 4%. The Thioflavin S (ThioS) core (red) contains densely packed amyloid fibrils. The toxic halo (brown) contains Type 2 oligomers and extends ≈ 50 microns from the edge of the ThioS core.
34
The radius of ThioS cores (r) ranges from ≈ 10 to 25 microns. The ratio of R/r ranges from ≈ 3 to 6. When the ThioS plaque load equals r3/R3, the toxic halos in the brain become confluent. This occurs when the ThioS plaque load is 1/63 to 1/33 = ≈ 0.5 to 4%. In humans with Alzheimer's disease, the ThioS plaque load ranges from ≈ 0.1% to 0.4%
37
References
-
- Vassar R, Bennett BD, Babu‐Khan S, et al. Beta‐secretase cleavage of Alzheimer's amyloid precursor protein by the transmembrane aspartic protease BACE. Science. 1999; 286(5440): 735‐741. - PubMed
-
- De Strooper B, Saftig P, Craessaerts K, et al. Deficiency of presenilin‐1 inhibits the normal cleavage of amyloid precursor protein. Nature. 1998; 391(6665): 387‐390. - PubMed
-
- Takasugi N, Tomita T, Hayashi I, et al. The role of presenilin cofactors in the gamma‐secretase complex. Nature. 2003; 422(6930): 438‐441. - PubMed
-
- Henley D, Raghavan N, Sperling R, Aisen P, Raman R, Romano G. Preliminary results of a trial of atabecestat in preclinical Alzheimer's disease. N Engl J Med. 2019; 380(15): 1483‐1485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
