

Chromosomal Evolution and Apomixis in the Cruciferous Tribe Boechereae
- PMID: 32547569
- PMCID: PMC7270200
- DOI: 10.3389/fpls.2020.00514
Chromosomal Evolution and Apomixis in the Cruciferous Tribe Boechereae
Abstract
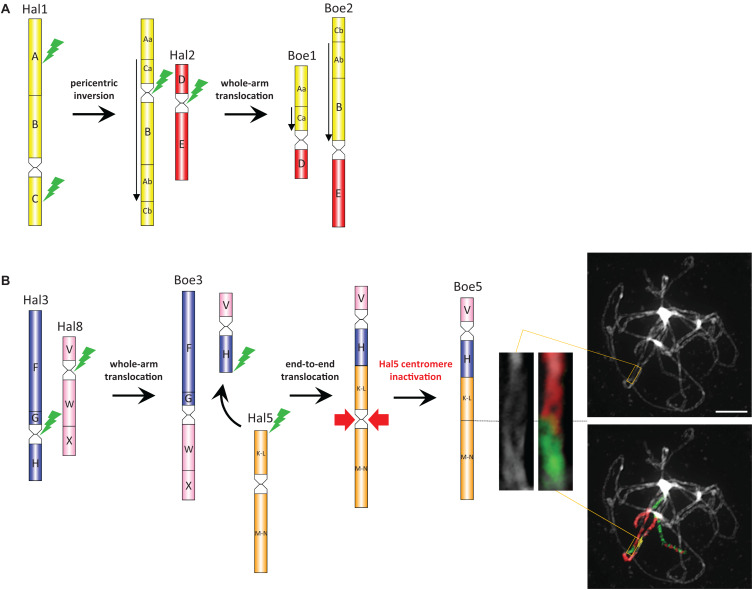
The mustard family (Brassicaceae) comprises several dozen monophyletic clades usually ranked as tribes. The tribe Boechereae plays a prominent role in plant research due to the incidence of apomixis and its close relationship to Arabidopsis. This tribe, largely confined to western North America, harbors nine genera and c. 130 species, with >90% of species belonging to the genus Boechera. Hundreds of apomictic diploid and triploid Boechera hybrids have spurred interest in this genus, but the remaining Boechereae genomes remain virtually unstudied. Here we report on comparative genome structure of six genera (Borodinia, Cusickiella, Phoenicaulis, Polyctenium, Nevada, and Sandbergia) and three Boechera species as revealed by comparative chromosome painting (CCP). All analyzed taxa shared the same seven-chromosome genome structure. Comparisons with the sister Halimolobeae tribe (n = 8) showed that the ancestral Boechereae genome (n = 7) was derived from an older n = 8 genome by descending dysploidy followed by the divergence of extant Boechereae taxa. As tribal divergence post-dated the origin of four tribe-specific chromosomes, it is proposed that these chromosomal rearrangements were a key evolutionary innovation underlaying the origin and diversification of the Boechereae in North America. Although most Boechereae genera exhibit genomic conservatism, intra-tribal cladogenesis has occasionally been accompanied by chromosomal rearrangements (particularly inversions). Recently, apomixis was reported in the Boechereae genera Borodinia and Phoenicaulis. Here, we report sexual reproduction in diploid Nevada, diploid Sandbergia, and tetraploid Cusickiella and aposporous apomixis in tetraploids of Polyctenium and Sandbergia. In sum, apomixis is now known to occur in five of the nine Boechereae genera.
Keywords: Cruciferae; North America; apomixis; apospory; autopolyploidy; descending dysploidy; karyotype evolution; speciation.
Copyright © 2020 Mandáková, Hloušková, Windham, Mitchell-Olds, Ashby, Price, Carman and Lysak.
Figures








References
-
- Alexander P. J., Windham M. D., Beck J. B., Al-Shehbaz I. A., Allphin L., Bailey C. D. (2013). Molecular phylogenetics and taxonomy of the genus Boechera and related genera (Brassicaceae: Boechereae). Syst. Bot. 38 192–209. 10.1600/036364413x661917 - DOI
-
- Alexander P. J., Windham M. D., Beck J. B., Al-Shehbaz I. A., Allphin L., Bailey C. D. (2015). Weaving a tangled web: divergent and reticulate speciation in Boecher fendleri sensu lato (Brassicaceae; Boechereae). Syst. Bot. 40 572–596. 10.1600/036364415x688745 - DOI
-
- Al-Shehbaz I. A. (2012). A generic and tribal synopsis of the Brassicaceae (Cruciferae). Taxon 61 931–954. 10.1002/tax.615002 - DOI
-
- Asker S. E., Jerling L. (1992). Apomixis in Plants. Boca Raton, FL: CRC Press.
-
- Bailey C. D., Al-Shehbaz I. A., Rajanikanth G. (2007). Generic limits in tribe Halimolobeae and description of the new genus Exhalimolobos (Brassicaceae). Syst. Bot. 32 140–156. 10.1600/036364407780360166 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials