

Mitochondria in cancer
- PMID: 32548570
- PMCID: PMC7278520
- DOI: 10.15698/cst2020.06.221
Mitochondria in cancer
Abstract
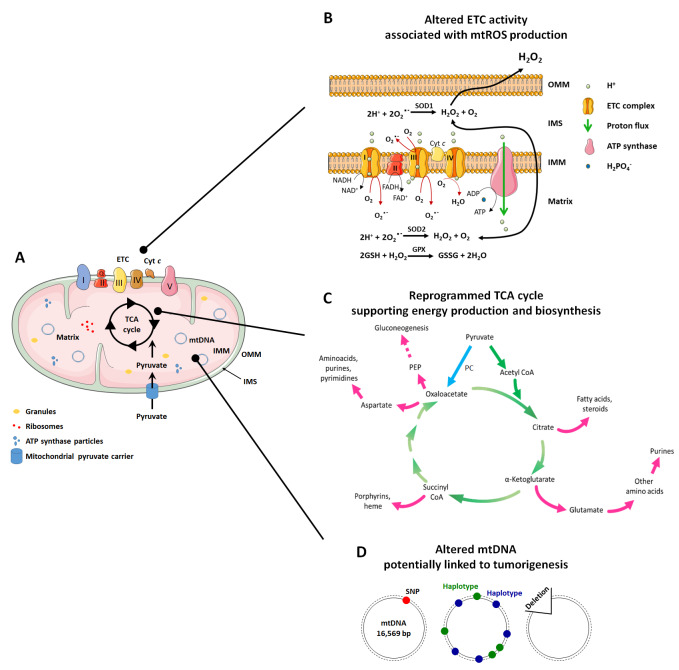
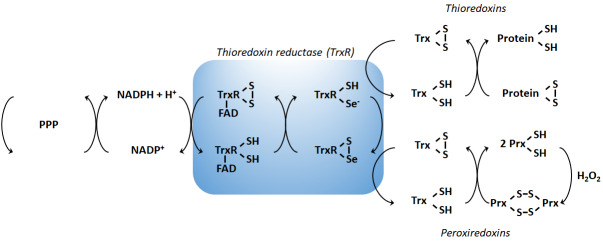
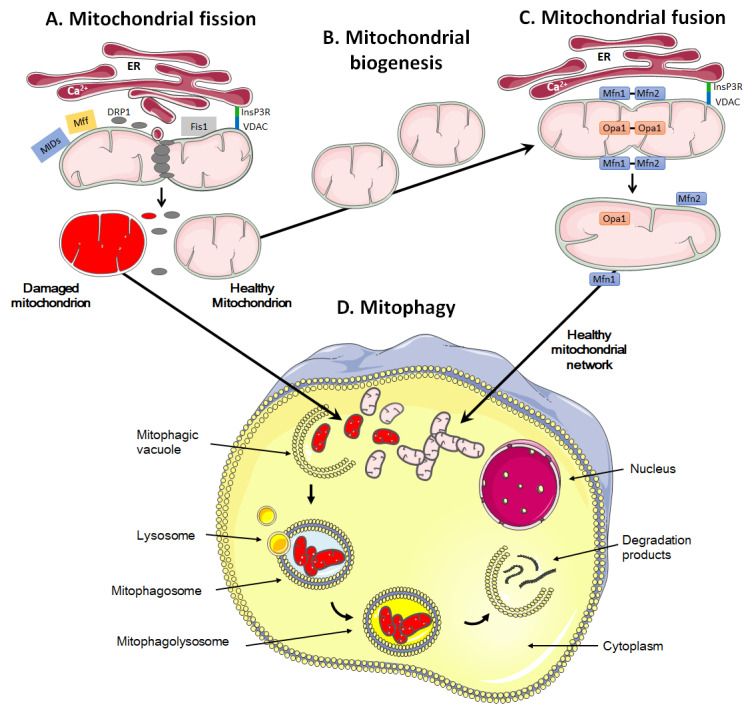
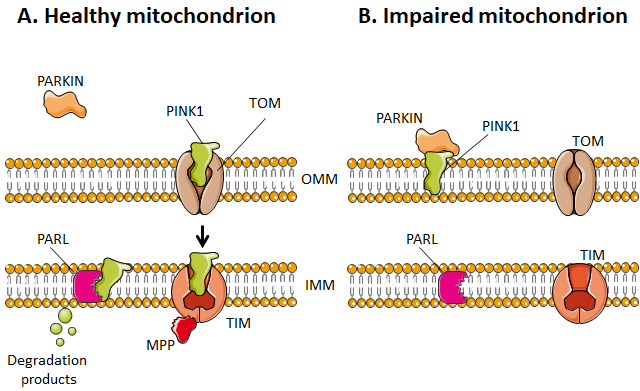
The rediscovery and reinterpretation of the Warburg effect in the year 2000 occulted for almost a decade the key functions exerted by mitochondria in cancer cells. Until recent times, the scientific community indeed focused on constitutive glycolysis as a hallmark of cancer cells, which it is not, largely ignoring the contribution of mitochondria to the malignancy of oxidative and glycolytic cancer cells, being Warburgian or merely adapted to hypoxia. In this review, we highlight that mitochondria are not only powerhouses in some cancer cells, but also dynamic regulators of life, death, proliferation, motion and stemness in other types of cancer cells. Similar to the cells that host them, mitochondria are capable to adapt to tumoral conditions, and probably to evolve to 'oncogenic mitochondria' capable of transferring malignant capacities to recipient cells. In the wider quest of metabolic modulators of cancer, treatments have already been identified targeting mitochondria in cancer cells, but the field is still in infancy.
Keywords: apoptosis; mitochondrial biogenesis; mitophagy; oxidative phosphorylation (OXPHOS); reactive oxygen species (ROS); tricarboxylic acid (TCA) cycle; tumor metabolism.
Copyright: © 2020 Grasso et al.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures
References
-
- Nomoto S, Yamashita K, Koshikawa K, Nakao A, Sidransky D. Mitochondrial D-loop mutations as clonal markers in multicentric hepatocellular carcinoma and plasma. Clin Cancer Res. 2002;8(2):481–487. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources