

The polycomb proteins EZH1 and EZH2 co-regulate chromatin accessibility and nephron progenitor cell lifespan in mice
- PMID: 32554463
- PMCID: PMC7450110
- DOI: 10.1074/jbc.RA120.013348
The polycomb proteins EZH1 and EZH2 co-regulate chromatin accessibility and nephron progenitor cell lifespan in mice
Abstract
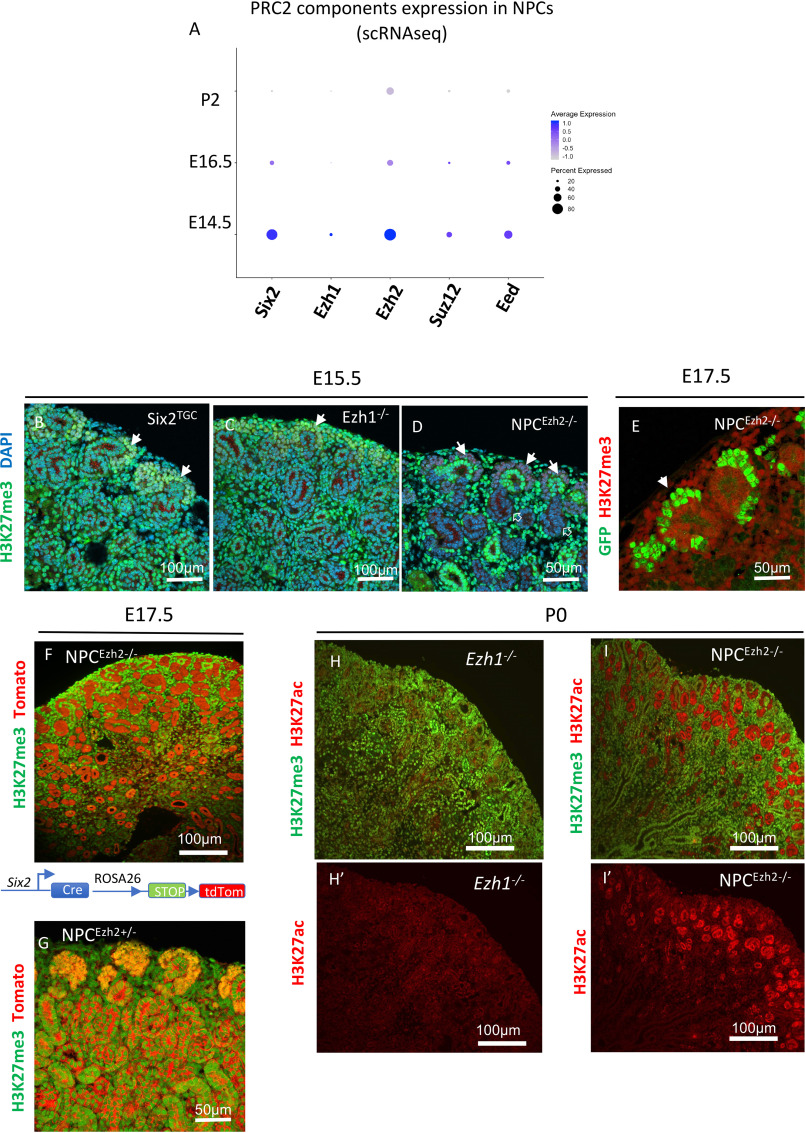
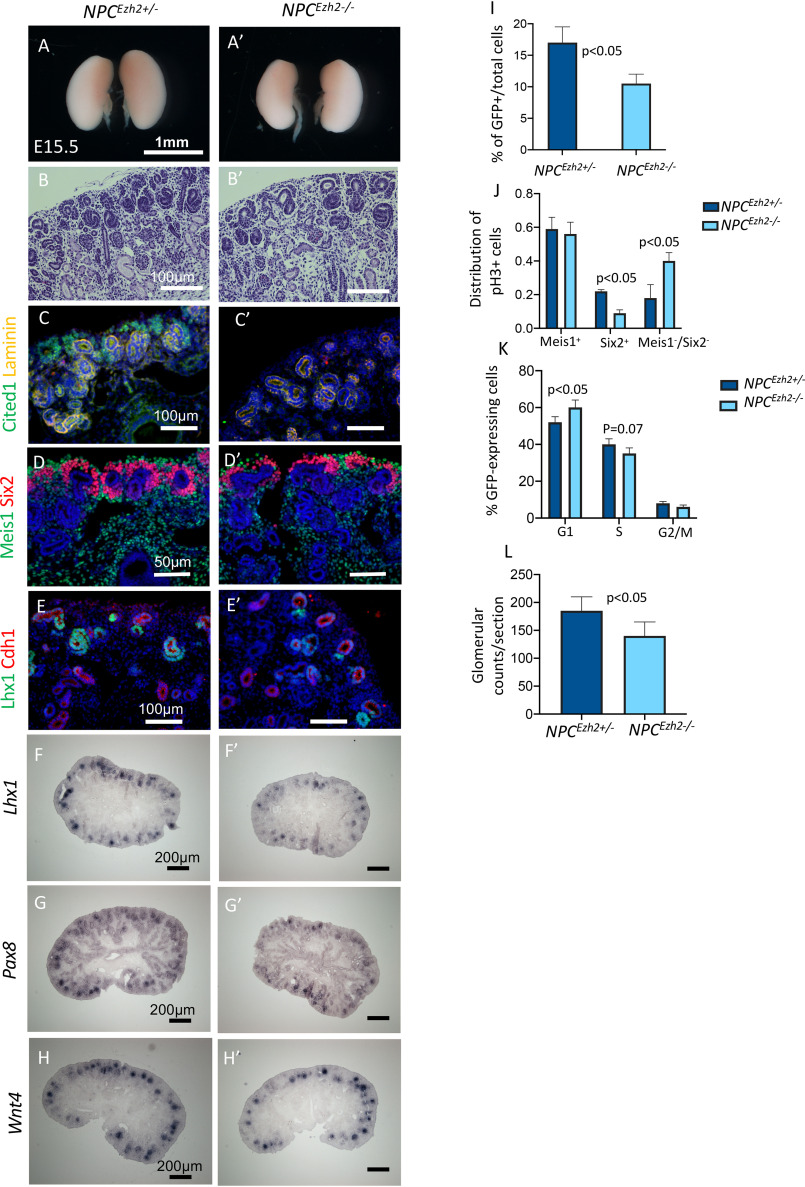
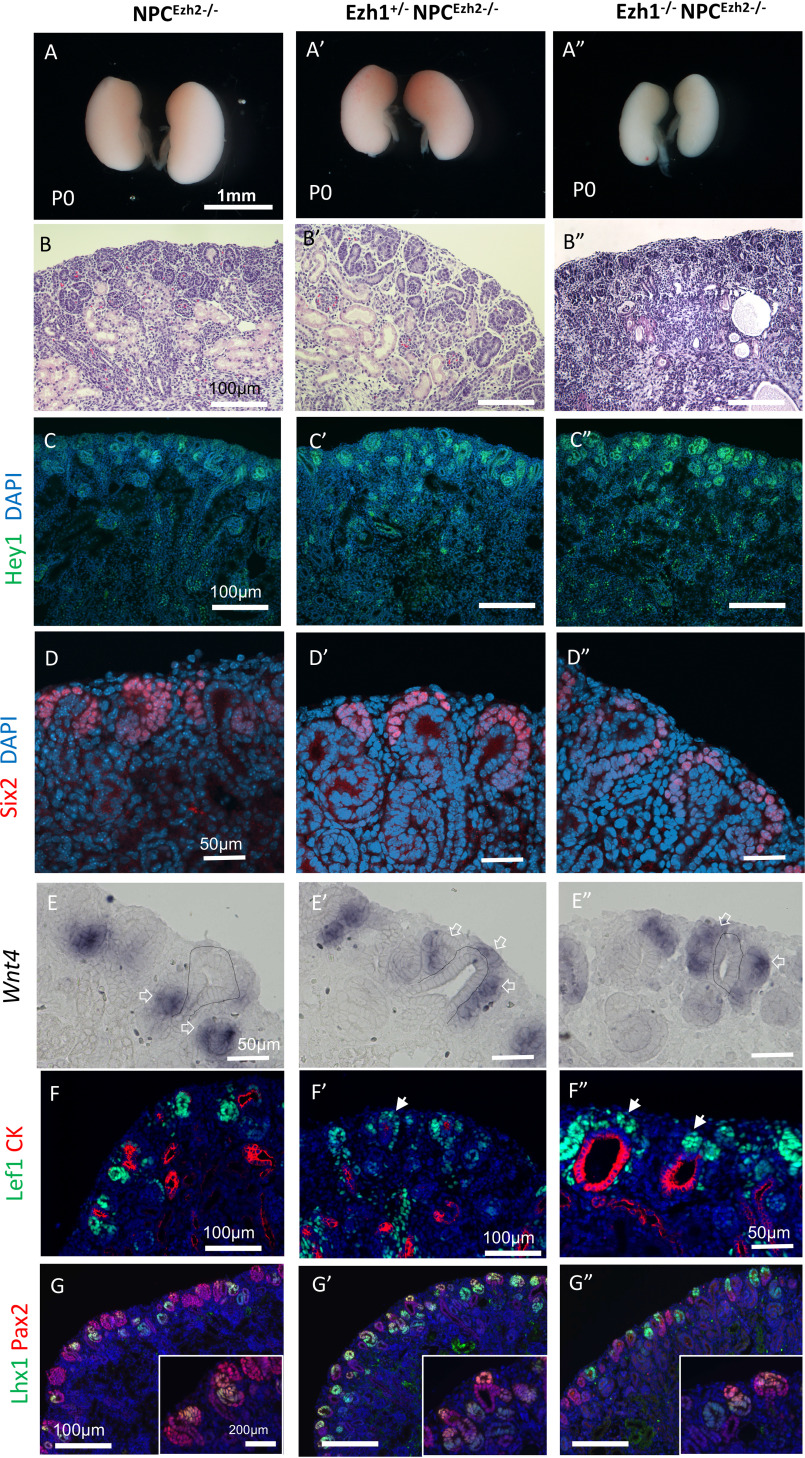
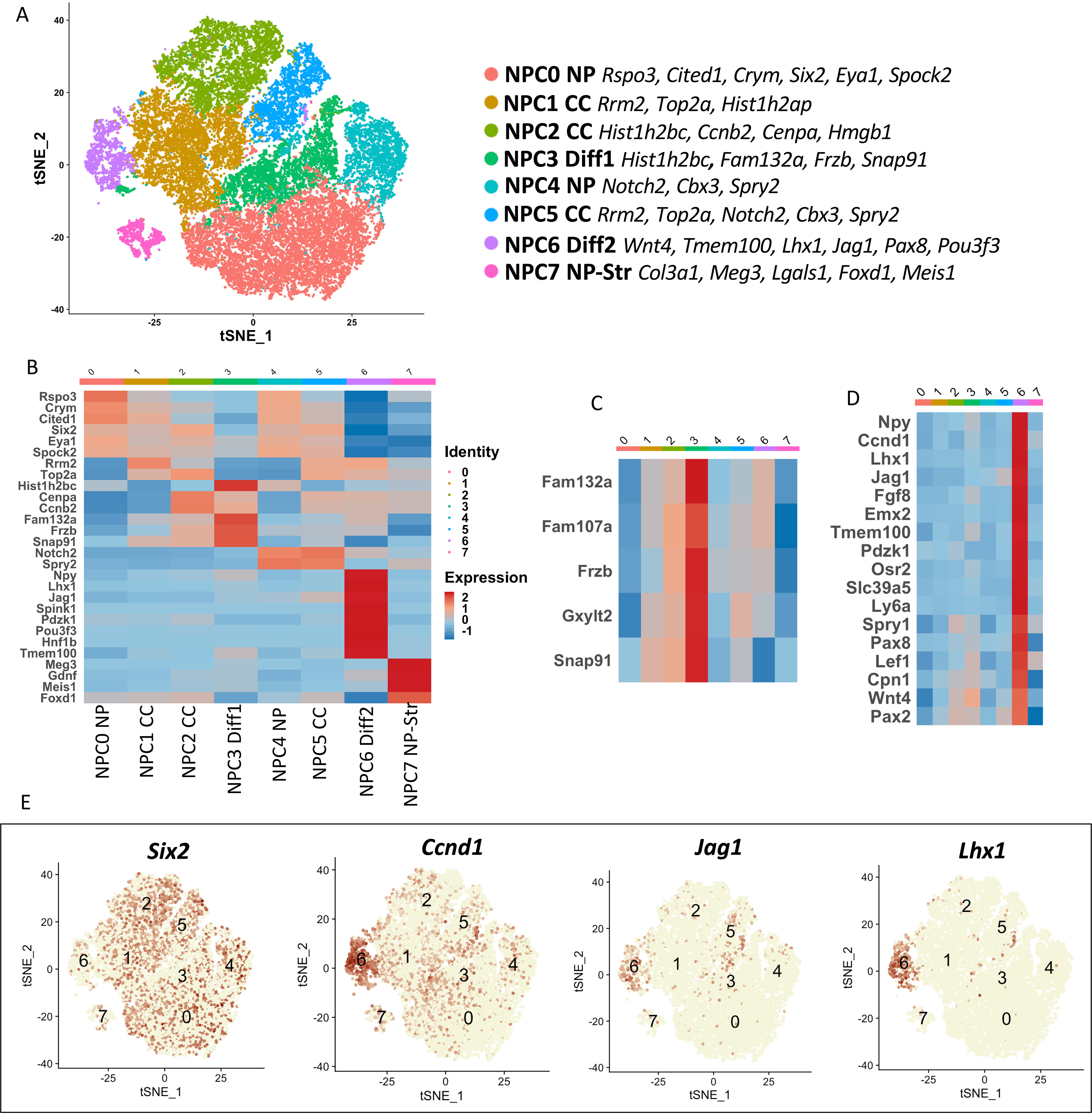
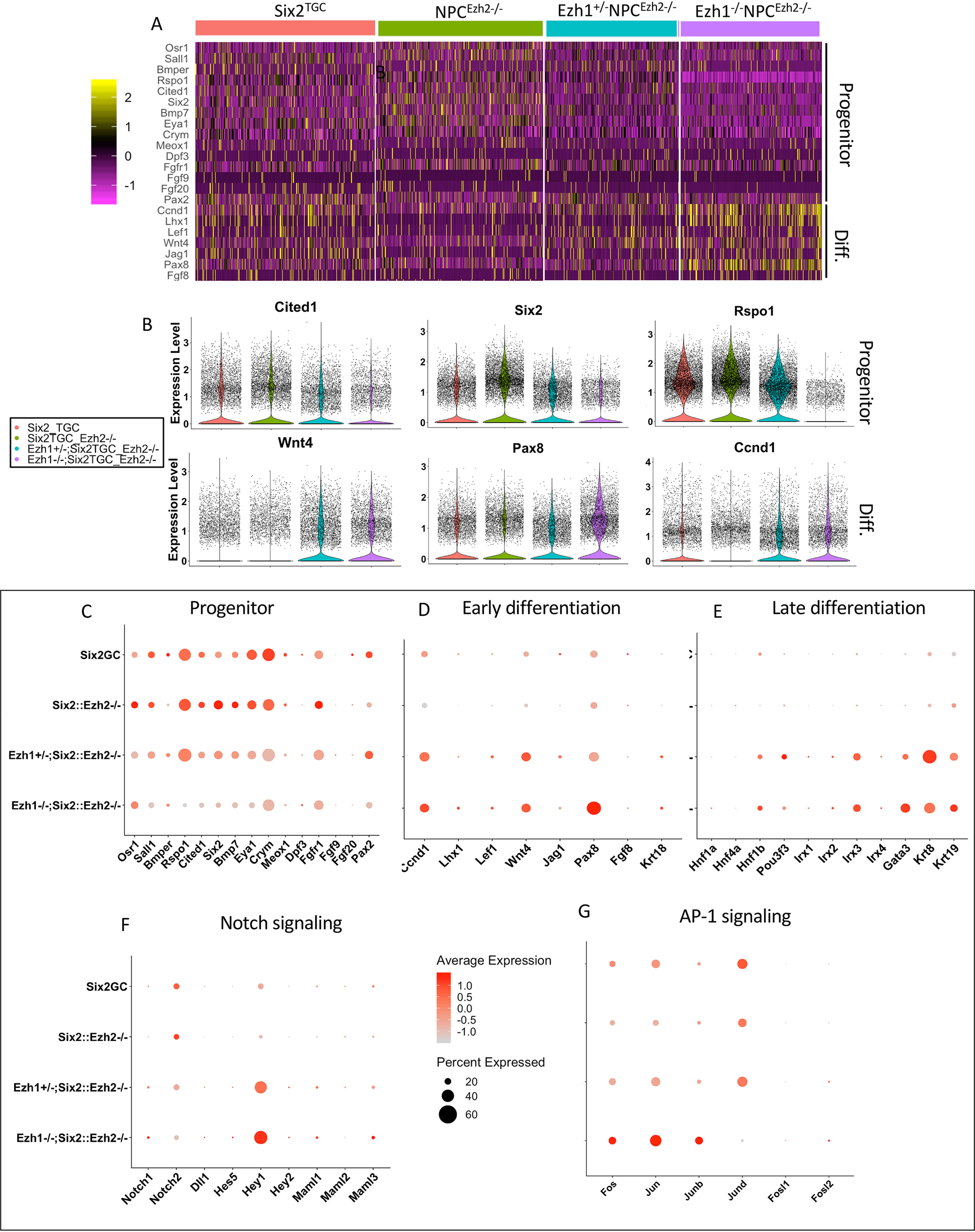
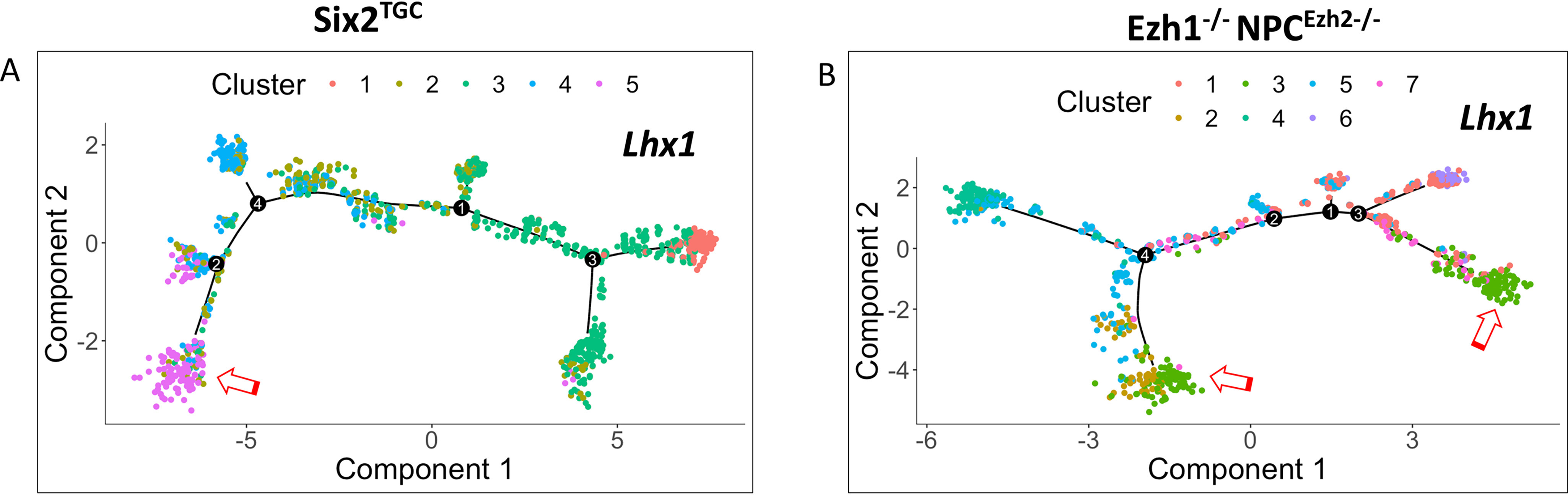
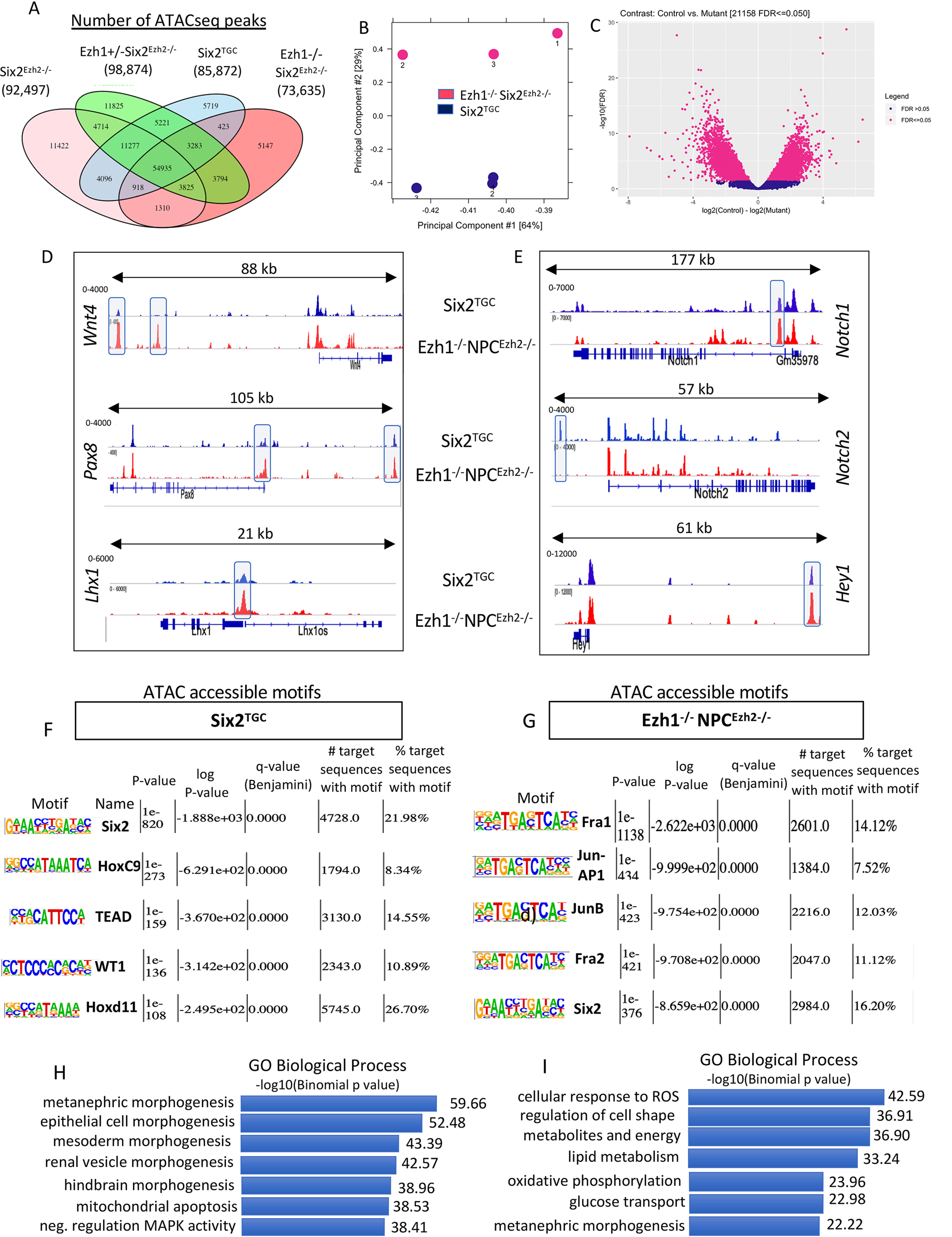
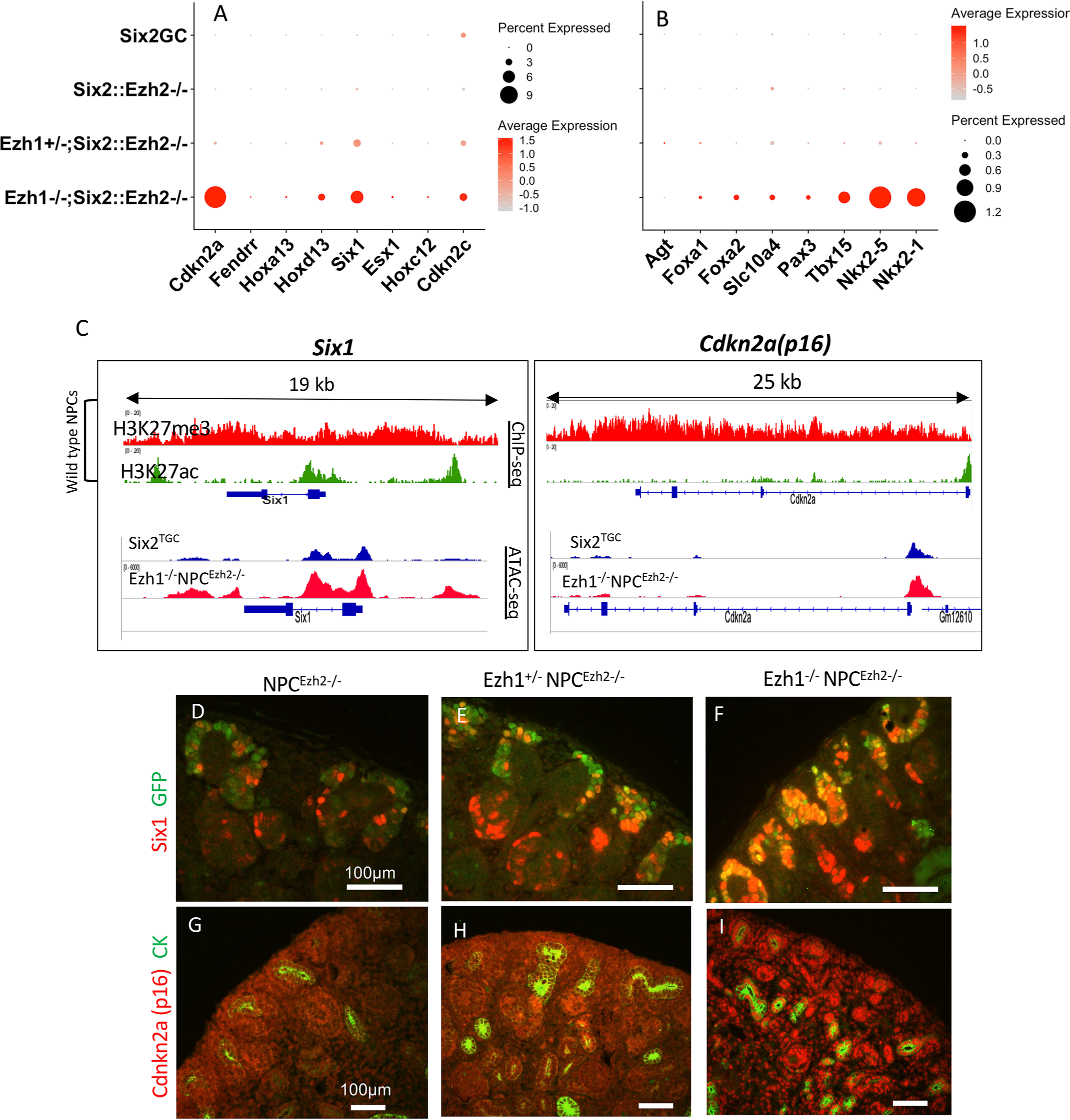
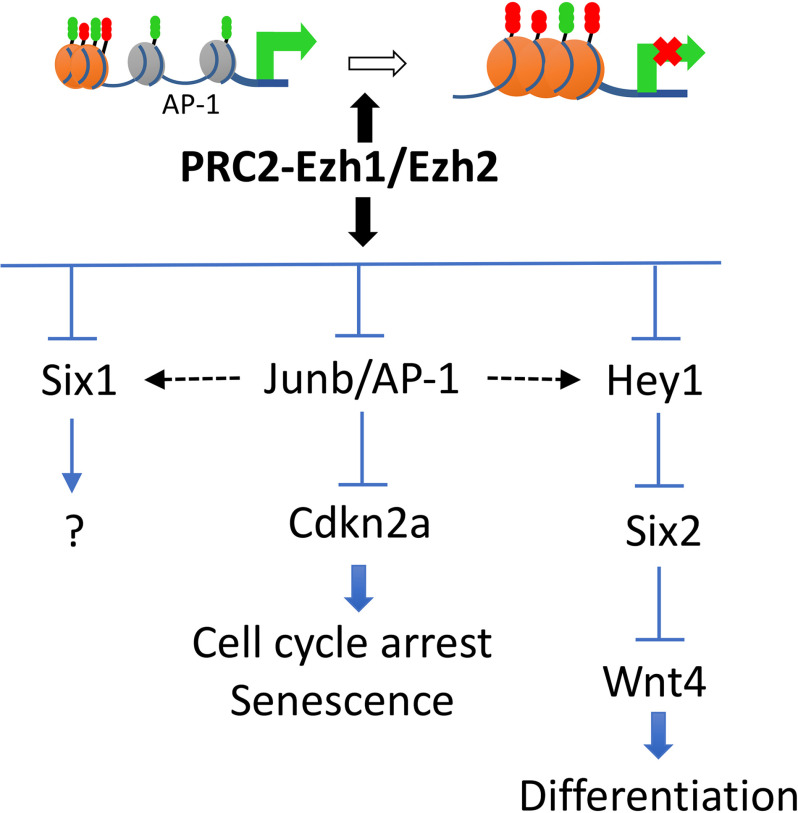
SIX2 (SIX homeobox 2)-positive nephron progenitor cells (NPCs) give rise to all epithelial cell types of the nephron, the filtering unit of the kidney. NPCs have a limited lifespan and are depleted near the time of birth. Epigenetic factors are implicated in the maintenance of organ-restricted progenitors such as NPCs, but the chromatin-based mechanisms are incompletely understood. Here, using a combination of gene targeting, chromatin profiling, and single-cell RNA analysis, we examined the role of the murine histone 3 Lys-27 (H3K27) methyltransferases EZH1 (enhancer of zeste 1) and EZH2 in NPC maintenance. We found that EZH2 expression correlates with NPC growth potential and that EZH2 is the dominant H3K27 methyltransferase in NPCs and epithelial descendants. Surprisingly, NPCs lacking H3K27 trimethylation maintained their progenitor state but cycled slowly, leading to a smaller NPC pool and formation of fewer nephrons. Unlike Ezh2 loss of function, dual inactivation of Ezh1 and Ezh2 triggered overexpression of the transcriptional repressor Hes-related family BHLH transcription factor with YRPW motif 1 (Hey1), down-regulation of Six2, and unscheduled activation of Wnt4-driven differentiation, resulting in early termination of nephrogenesis and severe renal dysgenesis. Double-mutant NPCs also overexpressed the SIX family member Six1 However, in this context, SIX1 failed to maintain NPC stemness. At the chromatin level, EZH1 and EZH2 restricted accessibility to AP-1-binding motifs, and their absence promoted a regulatory landscape akin to differentiated and nonlineage cells. We conclude that EZH2 is required for NPC renewal potential and that tempering of the differentiation program requires cooperation of both EZH1 and EZH2.
Keywords: H3K27 methyltransferase; SIX homeobox 2 (SIX2); chromatin; development; enhancer of zeste (Ezh); epigenetics; histone methylation; kidney; nephrology; nephron progenitors; polycomb.
© 2020 Liu et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Kobayashi A., Valerius M. T., Mugford J. W., Carroll T. J., Self M., Oliver G., and McMahon A. P. (2008) Six2 defines and regulates a multipotent self-renewing nephron progenitor population throughout mammalian kidney development. Cell Stem Cell 3, 169–181 10.1016/j.stem.2008.05.020 - DOI - PMC - PubMed
-
- Lindström N. O., McMahon J. A., Guo J., Tran T., Guo Q., Rutledge E., Parvez R. K., Saribekyan G., Schuler R. E., Liao C., Kim A. D., Abdelhalim A., Ruffins S. W., Thornton M. E., Basking L., et al. (2018) Conserved and divergent features of human and mouse kidney organogenesis. J. Am. Soc. Nephrol. 29, 785–805 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
