

Strategies to Modulate MicroRNA Functions for the Treatment of Cancer or Organ Injury
- PMID: 32554488
- PMCID: PMC7300323
- DOI: 10.1124/pr.119.019026
Strategies to Modulate MicroRNA Functions for the Treatment of Cancer or Organ Injury
Abstract
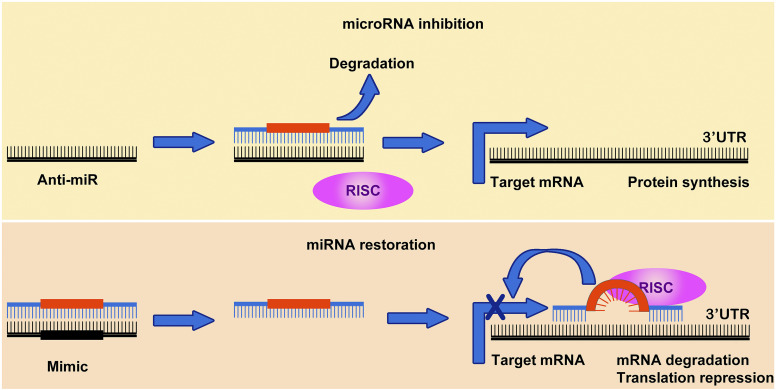
Cancer and organ injury-such as that occurring in the perioperative period, including acute lung injury, myocardial infarction, and acute gut injury-are among the leading causes of death in the United States and impose a significant impact on quality of life. MicroRNAs (miRNAs) have been studied extensively during the last two decades for their role as regulators of gene expression, their translational application as diagnostic markers, and their potential as therapeutic targets for disease treatment. Despite promising preclinical outcomes implicating miRNA targets in disease treatment, only a few miRNAs have reached clinical trials. This likely relates to difficulties in the delivery of miRNA drugs to their targets to achieve efficient inhibition or overexpression. Therefore, understanding how to efficiently deliver miRNAs into diseased tissues and specific cell types in patients is critical. This review summarizes current knowledge on various approaches to deliver therapeutic miRNAs or miRNA inhibitors and highlights current progress in miRNA-based disease therapy that has reached clinical trials. Based on ongoing advances in miRNA delivery, we believe that additional therapeutic approaches to modulate miRNA function will soon enter routine medical treatment of human disease, particularly for cancer or perioperative organ injury. SIGNIFICANCE STATEMENT: MicroRNAs have been studied extensively during the last two decades in cancer and organ injury, including acute lung injury, myocardial infarction, and acute gut injury, for their regulation of gene expression, application as diagnostic markers, and therapeutic potentials. In this review, we specifically emphasize the pros and cons of different delivery approaches to modulate microRNAs, as well as the most recent exciting progress in the field of therapeutic targeting of microRNAs for disease treatment in patients.
Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics.
Figures
References
-
- Abdelmawla S, Guo S, Zhang L, Pulukuri SM, Patankar P, Conley P, Trebley J, Guo P, Li QX. (2011) Pharmacological characterization of chemically synthesized monomeric phi29 pRNA nanoparticles for systemic delivery [published correction appears in Mol Ther (2011) 19:1748]. Mol Ther 19:1312–1322. - PMC - PubMed
-
- Adams D, Gonzalez-Duarte A, O’Riordan WD, Yang CC, Ueda M, Kristen AV, Tournev I, Schmidt HH, Coelho T, Berk JL, et al. (2018) Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N Engl J Med 379:11–21. - PubMed
-
- Anderson JL, Morrow DA. (2017) Acute myocardial infarction. N Engl J Med 376:2053–2064. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA150153/CA/NCI NIH HHS/United States
- R01 NS064607/NS/NINDS NIH HHS/United States
- P01 HL114457/HL/NHLBI NIH HHS/United States
- R01 HL109233/HL/NHLBI NIH HHS/United States
- UL1 TR003167/TR/NCATS NIH HHS/United States
- P30 DK056338/DK/NIDDK NIH HHS/United States
- R01 DK097075/DK/NIDDK NIH HHS/United States
- R01 NS104280/NS/NINDS NIH HHS/United States
- R01 DK109574/DK/NIDDK NIH HHS/United States
- P01 CA163205/CA/NCI NIH HHS/United States
- R01 HL119837/HL/NHLBI NIH HHS/United States
- R01 HL133900/HL/NHLBI NIH HHS/United States
- R01 DK122796/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
