

Molecular approaches underlying the oogenic cycle of the scleractinian coral, Acropora tenuis
- PMID: 32555307
- PMCID: PMC7303178
- DOI: 10.1038/s41598-020-66020-x
Molecular approaches underlying the oogenic cycle of the scleractinian coral, Acropora tenuis
Abstract
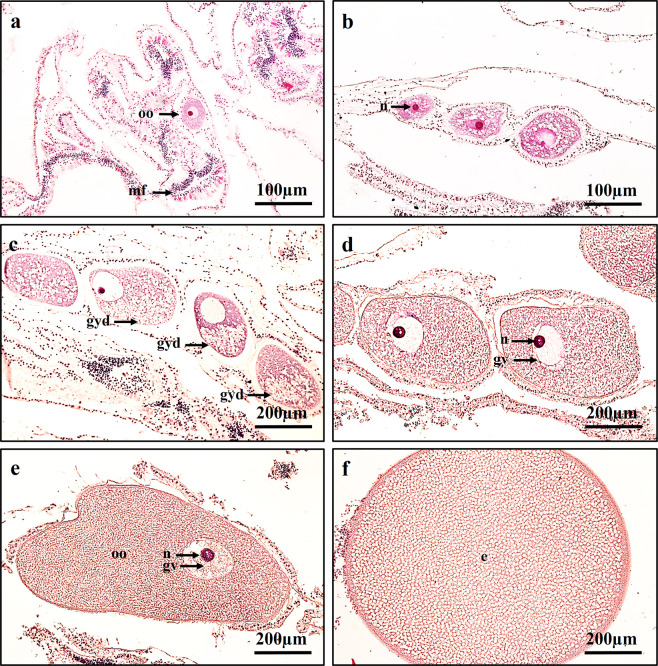
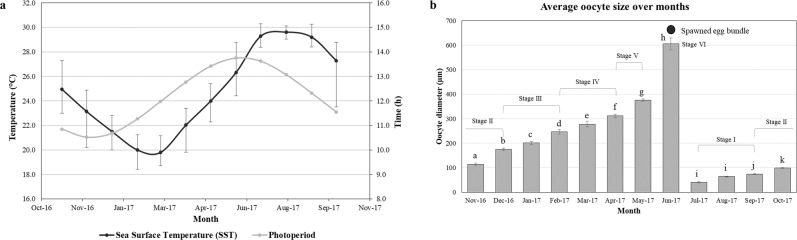
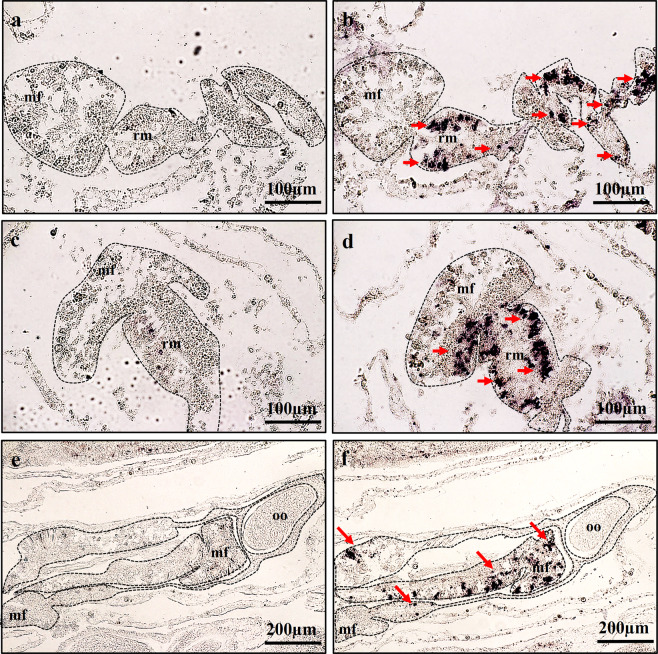
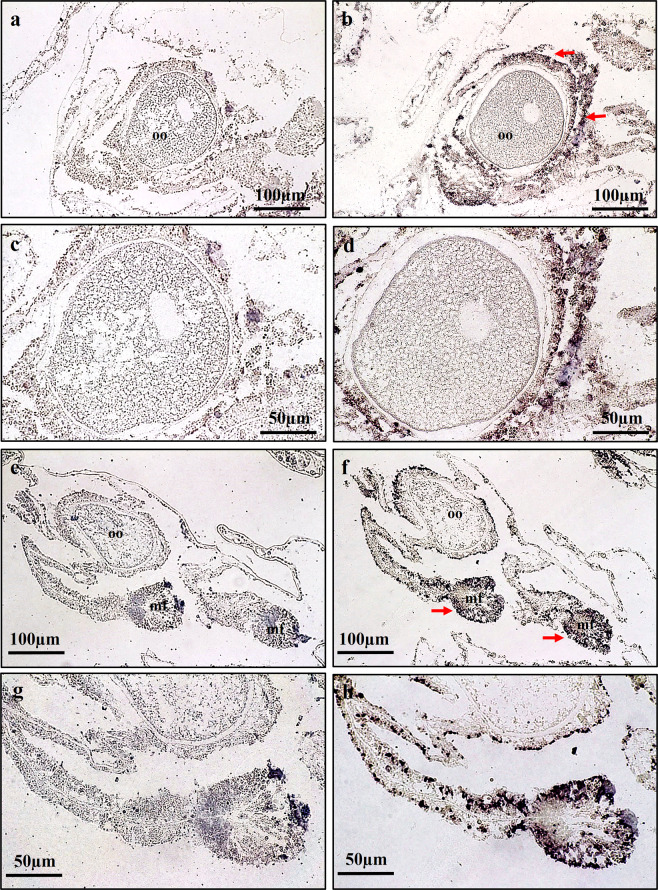
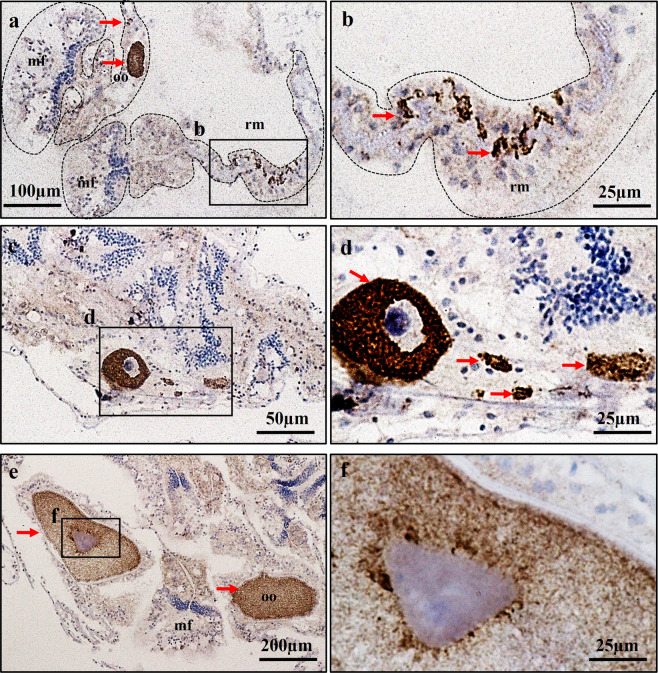
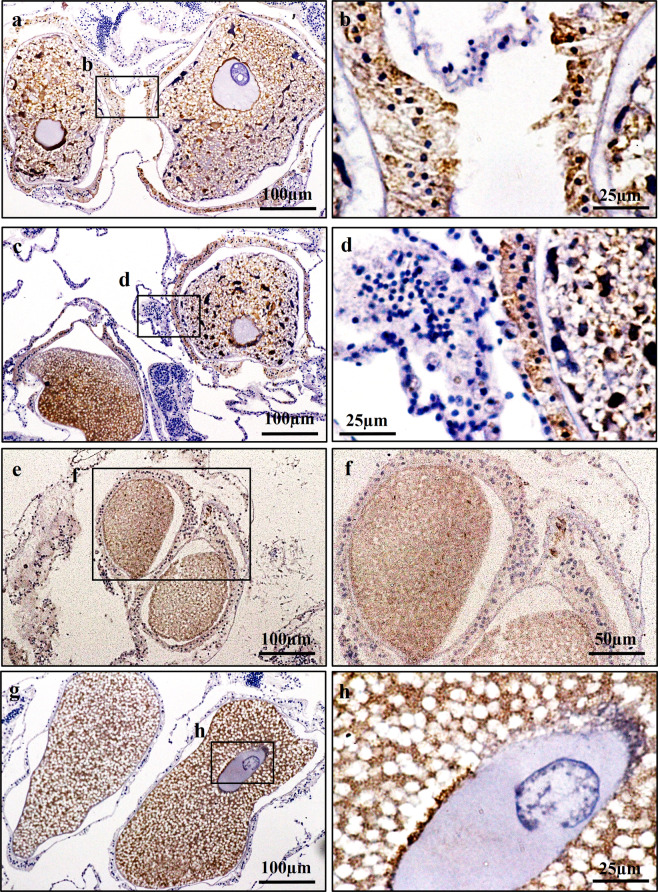
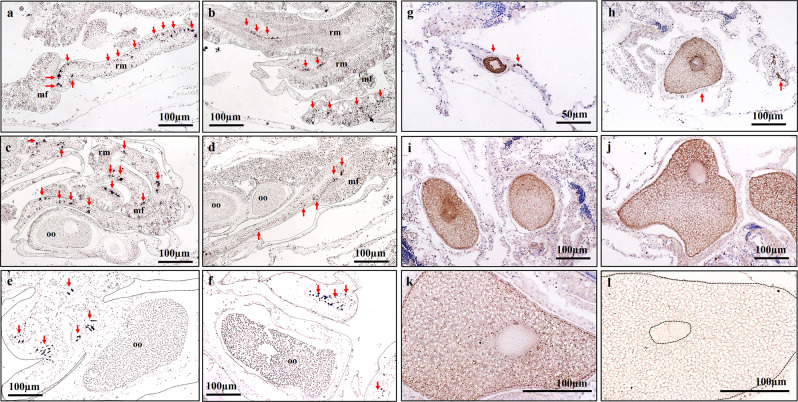
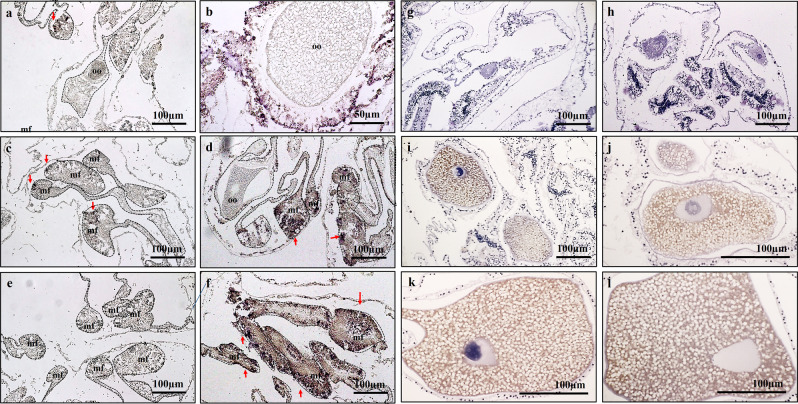
This study aimed to elucidate the physiological processes of oogenesis in Acropora tenuis. Genes/proteins related to oogenesis were investigated: Vasa, a germ cell marker, vitellogenin (VG), a major yolk protein precursor, and its receptor (LDLR). Coral branches were collected monthly from coral reefs around Sesoko Island (Okinawa, Japan) for histological observation by in situ hybridisation (ISH) of the Vasa (AtVasa) and Low Density Lipoprotein Receptor (AtLDLR) genes and immunohistochemistry (IHC) of AtVasa and AtVG. AtVasa immunoreactivity was detected in germline cells and ooplasm, whereas AtVG immunoreactivity was detected in ooplasm and putative ovarian tissues. AtVasa was localised in germline cells located in the retractor muscles of the mesentery, whereas AtLDLR was localised in the putative ovarian and mesentery tissues. AtLDLR was detected in coral tissues during the vitellogenic phase, whereas AtVG immunoreactivity was found in primary oocytes. Germline cells expressing AtVasa are present throughout the year. In conclusion, Vasa has physiological and molecular roles throughout the oogenic cycle, as it determines gonadal germline cells and ensures normal oocyte development, whereas the roles of VG and LDLR are limited to the vitellogenic stages because they act in coordination with lipoprotein transport, vitellogenin synthesis, and yolk incorporation into oocytes.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Chui PY, et al. Gametogenesis, Embryogenesis, and Fertilization Ecology of Platygyra acuta in Marginal Nonreefal Coral Communities in Hong Kong. J. Mar. Biol. 2014;2014:1–9.
-
- Harrison, P. L. Sexual reproduction in scleractinian corals in Coral reefs: an ecosystem in transition (eds. Dubinsky, Z., Stambler, N.) 59-85 (Springer, Dordrecht, 2011).
-
- Harrison, P. L., Wallace, C. C. Reproduction, dispersal and recruitment of scleractinian coral in Ecosystem of the World Vol. 25 (ed. Dubinsky, Z.) 133–207 (Elsevier, Amsterdam, 1990).
-
- Isomura, N. & Fukami, H. Coral Reproduction in Japan in Coral Reef Studies of Japan (ed. Iguchi A.) 95–110 (Springer, Dordrecht, 2018).
-
- Harrison PL, et al. Mass spawning in tropical reef corals. Science. 1984;223:1186–1189. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
