

Senescent cholangiocytes release extracellular vesicles that alter target cell phenotype via the epidermal growth factor receptor
- PMID: 32558183
- PMCID: PMC7669612
- DOI: 10.1111/liv.14569
Senescent cholangiocytes release extracellular vesicles that alter target cell phenotype via the epidermal growth factor receptor
Abstract
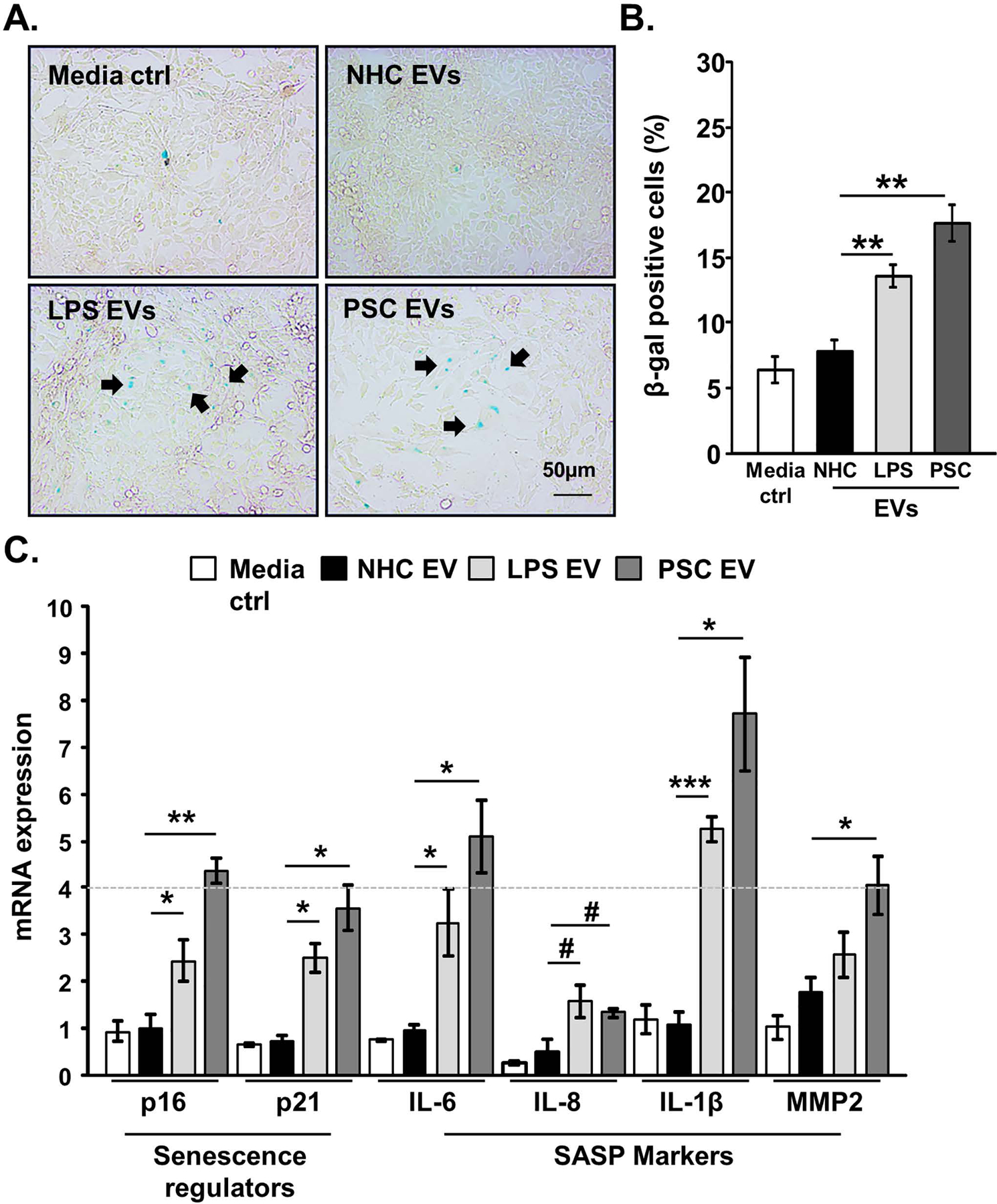
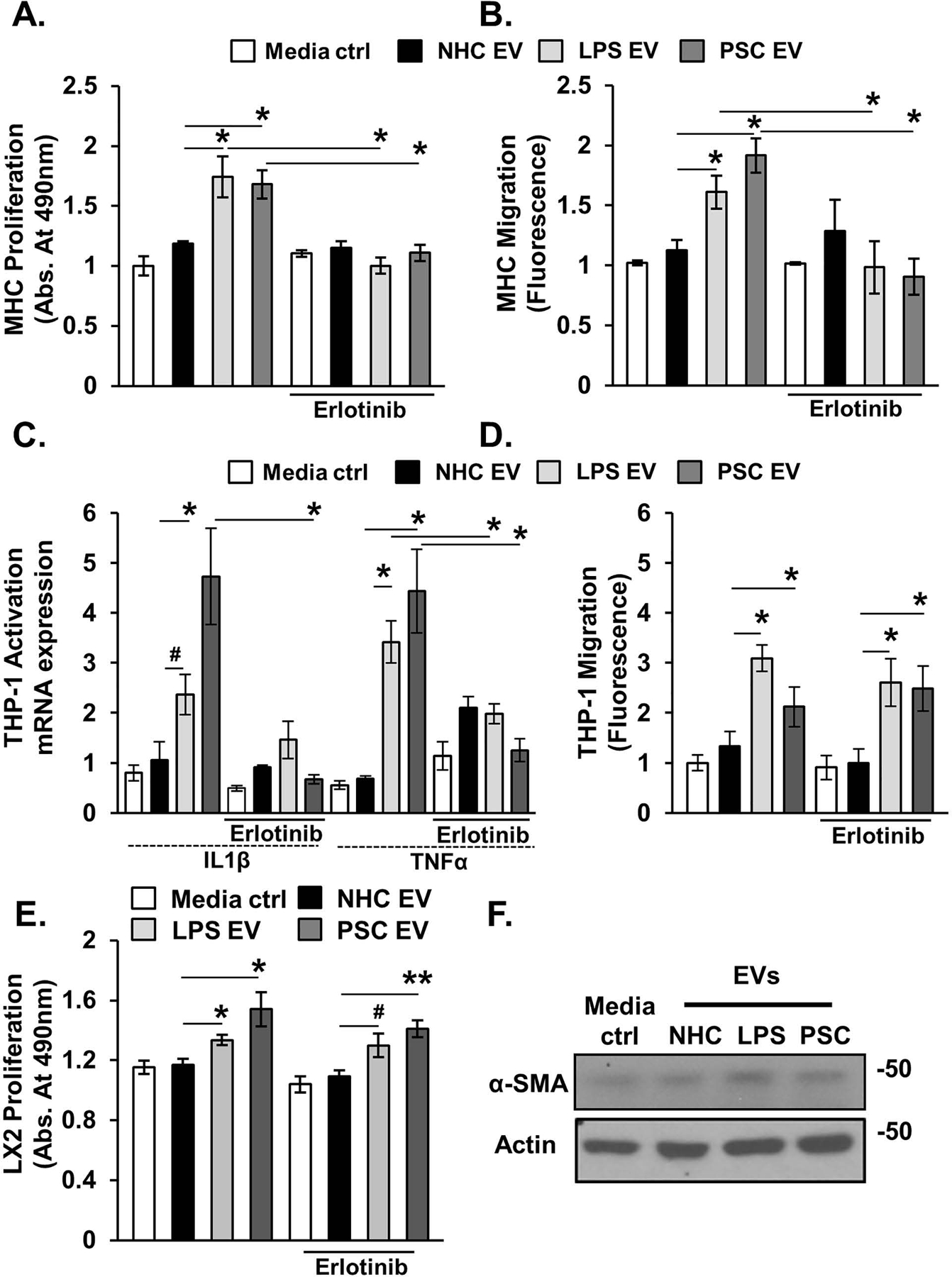
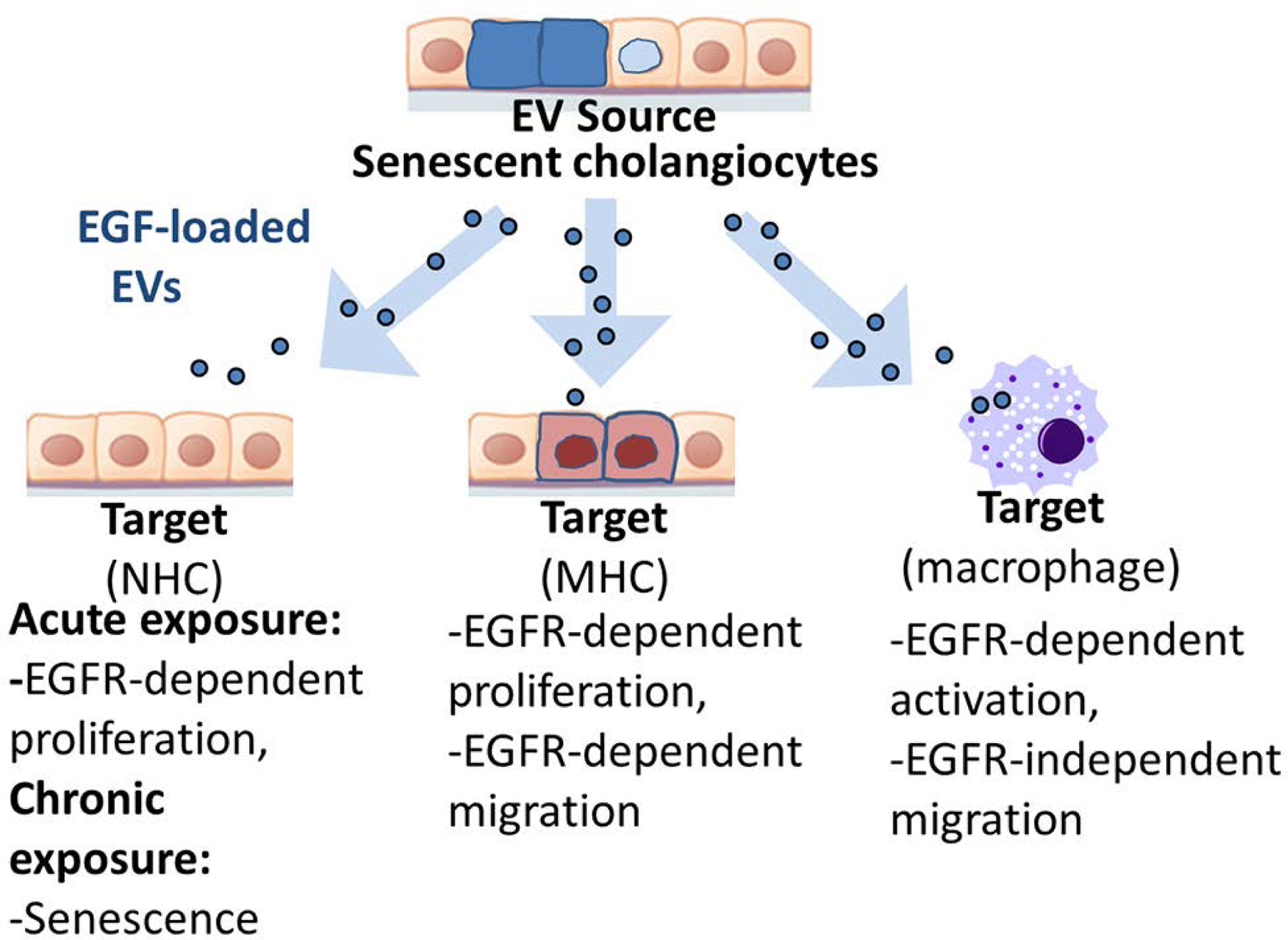
Background & aims: Primary sclerosing cholangitis (PSC) is a chronic liver disease characterized by peribiliary inflammation and fibrosis. Cholangiocyte senescence is a prominent feature of PSC. Here, we hypothesize that extracellular vesicles (EVs) from senescent cholangiocytes influence the phenotype of target cells.
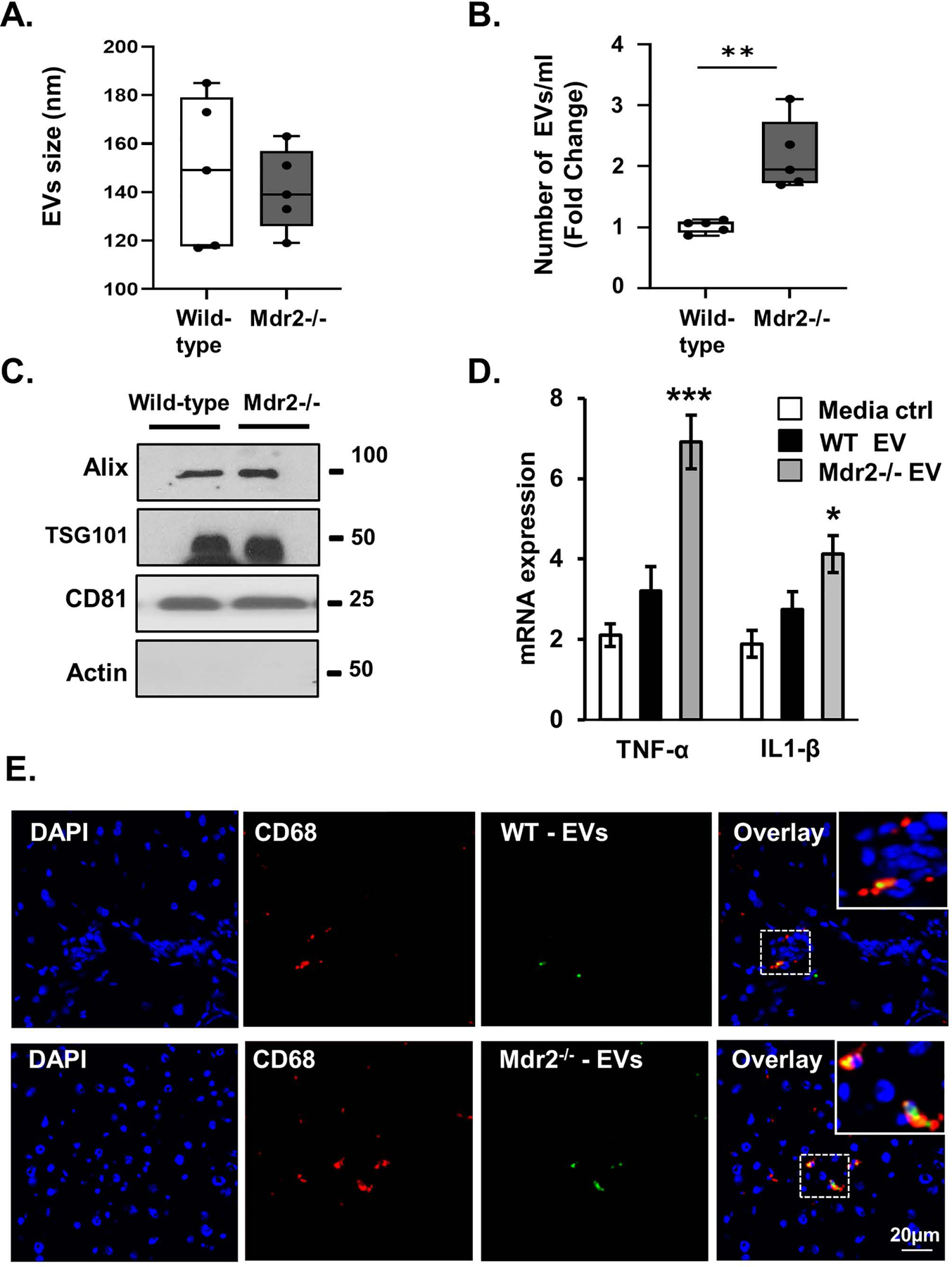
Methods: EVs were isolated from normal human cholangiocytes (NHCs), cholangiocytes from PSC patients and NHCs experimentally induced to senescence. NHCs, malignant human cholangiocytes (MHCs) and monocytes were exposed to 108 EVs from each donor cell population and assessed for proliferation, MAPK activation and migration. Additionally, we isolated EVs from plasma of wild-type and Mdr2-/- mice (a murine model of PSC), and assessed mouse monocyte activation.
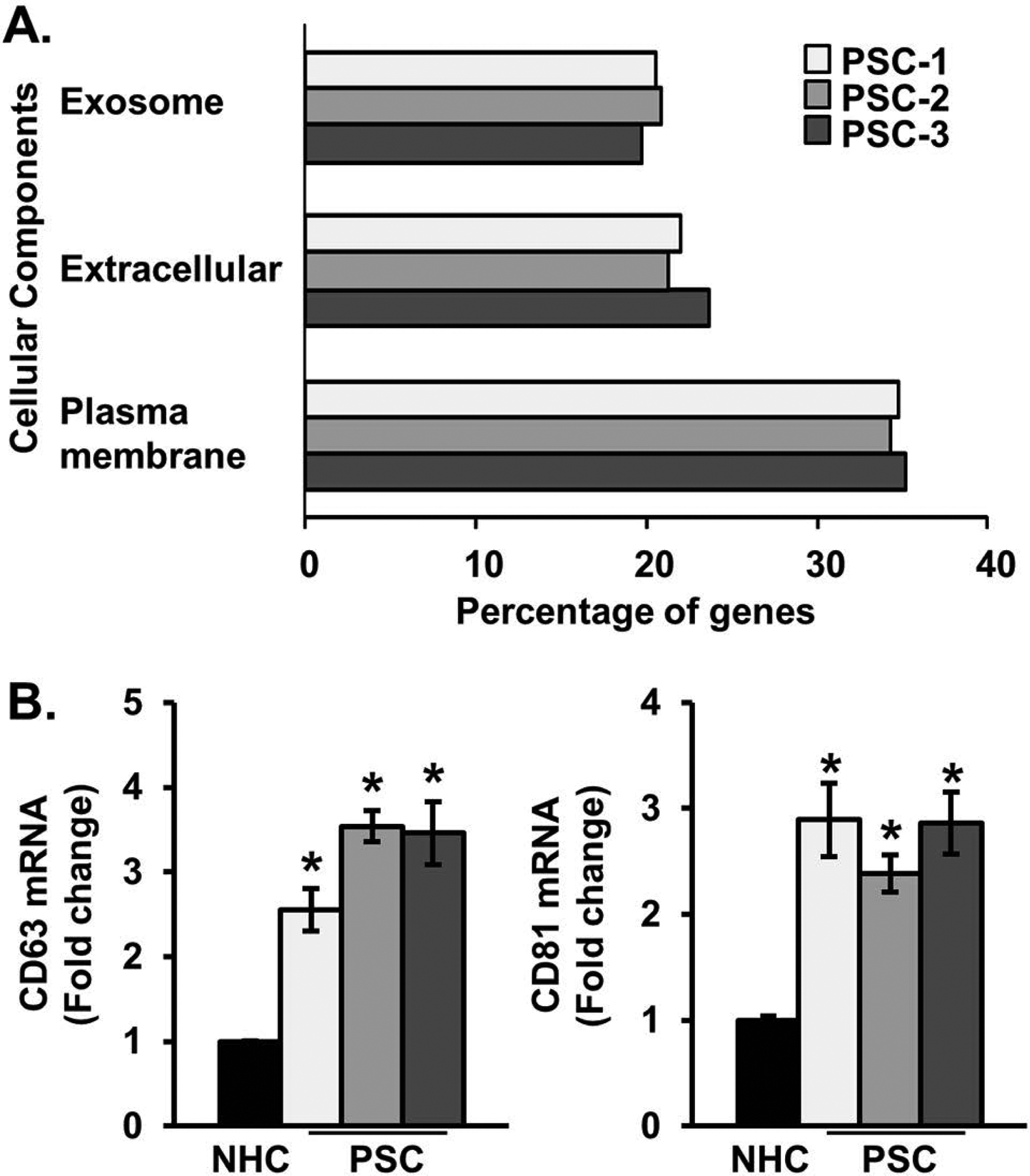
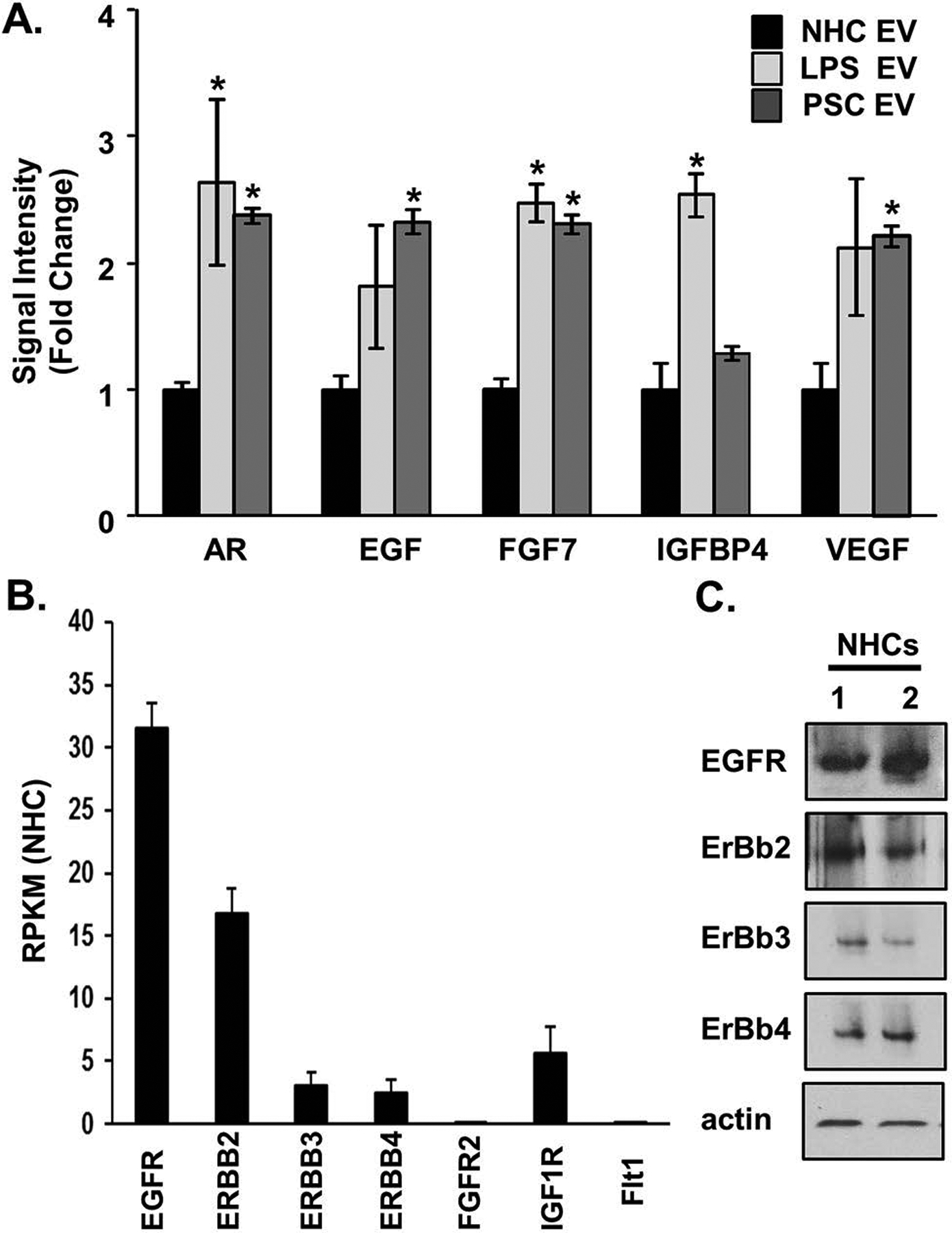
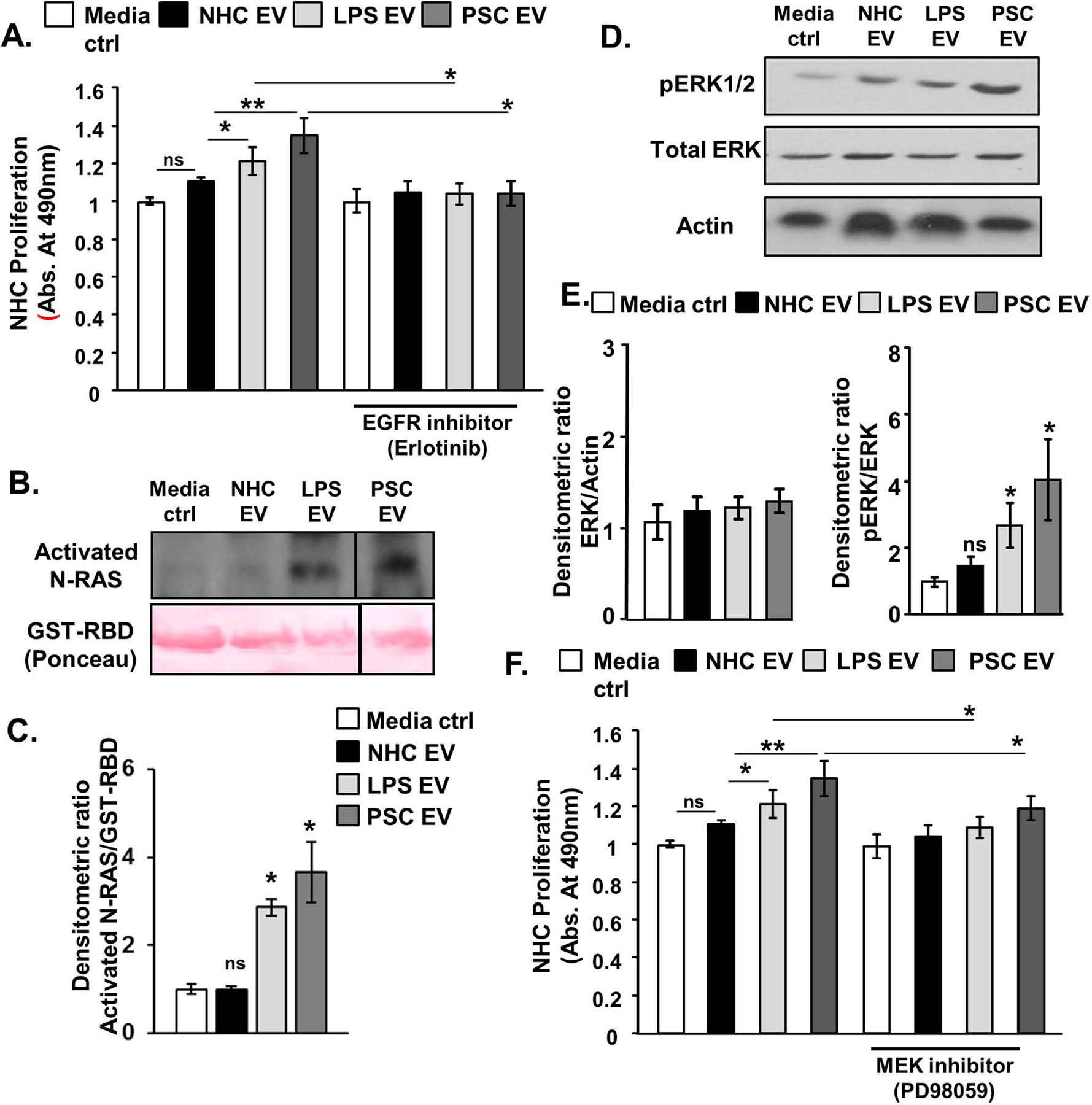
Results: EVs exhibited the size and protein markers of exosomes. The number of EVs released from senescent human cholangiocytes was increased; similarly, the EVs in plasma from Mdr2-/- mice were increased. Additionally, EVs from senescent cholangiocytes were enriched in multiple growth factors, including EGF. NHCs exposed to EVs from senescent cholangiocytes showed increased NRAS and ERK1/2 activation. Moreover, EVs from senescent cholangiocytes promoted proliferation of NHCs and MHCs, findings that were blocked by erlotinib, an EGF receptor inhibitor. Furthermore, EVs from senescent cholangiocytes induced EGF-dependent Interleukin 1-beta and Tumour necrosis factor expression and migration of human monocytes; similarly, Mdr2-/- mouse plasma EVs induced activation of mouse monocytes.
Conclusions: The data continue to support the importance of cholangiocyte senescence in PSC pathogenesis, directly implicate EVs in cholangiocyte proliferation, malignant progression and immune cell activation and migration, and identify novel therapeutic approaches for PSC.
Keywords: biliary epithelial cell; cellular senescence; extracellular vesicles; primary sclerosing cholangitis; senescence-associated secretory phenotype.
© 2020 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Conflict of interest statement
Figures

References
-
- Bashir MR, Muir AJ. Great Expectations - Can Magnetic Resonance Elastography Accelerate Progress in Primary Sclerosing Cholangitis Research? Clin Gastroenterol Hepatol 2019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
