

Direct Readout of Neural Stem Cell Transgenesis with an Integration-Coupled Gene Expression Switch
- PMID: 32559415
- PMCID: PMC7447981
- DOI: 10.1016/j.neuron.2020.05.038
Direct Readout of Neural Stem Cell Transgenesis with an Integration-Coupled Gene Expression Switch
Abstract
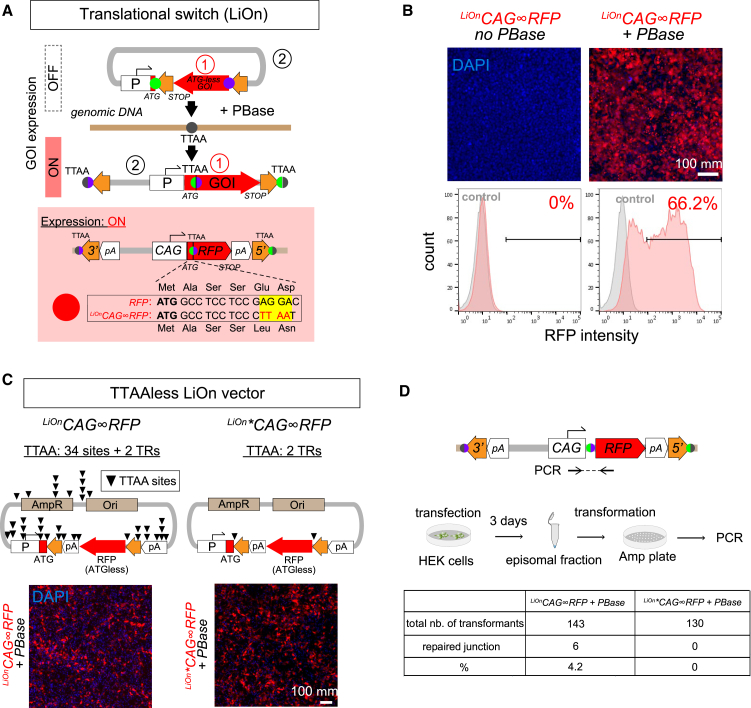
Stable genomic integration of exogenous transgenes is essential in neurodevelopmental and stem cell studies. Despite tools driving increasingly efficient genomic insertion with DNA vectors, transgenesis remains fundamentally hindered by the impossibility of distinguishing integrated from episomal transgenes. Here, we introduce an integration-coupled On genetic switch, iOn, which triggers gene expression upon incorporation into the host genome through transposition, thus enabling rapid and accurate identification of integration events following transfection with naked plasmids. In vitro, iOn permits rapid drug-free stable transgenesis of mouse and human pluripotent stem cells with multiple vectors. In vivo, we demonstrate faithful cell lineage tracing, assessment of regulatory elements, and mosaic analysis of gene function in somatic transgenesis experiments that reveal neural progenitor potentialities and interaction. These results establish iOn as a universally applicable strategy to accelerate and simplify genetic engineering in cultured systems and model organisms by conditioning transgene activation to genomic integration.
Keywords: DNA vectors; genetic engineering; genetic switch; genomic integration; lineage tracing; mosaic analysis; neural stem cells; somatic transgenesis; transposon systems.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests J.L., T.K., R.B.-M., F.M., S.T., K.L., and M. Le have filed a patent application related to the technology described in this article (number EP18305623).
Figures







References
-
- Akhtar W., de Jong J., Pindyurin A.V., Pagie L., Meuleman W., de Ridder J., Berns A., Wessels L.F.A., van Lohuizen M., van Steensel B. Chromatin position effects assayed by thousands of reporters integrated in parallel. Cell. 2013;154:914–927. - PubMed
-
- Batard P., Jordan M., Wurm F. Transfer of high copy number plasmid into mammalian cells by calcium phosphate transfection. Gene. 2001;270:61–68. - PubMed
-
- Beard C., Hochedlinger K., Plath K., Wutz A., Jaenisch R. Efficient method to generate single-copy transgenic mice by site-specific integration in embryonic stem cells. Genesis. 2006;44:23–28. - PubMed
-
- Black J.B., Perez-Pinera P., Gersbach C.A. Mammalian synthetic biology: engineering biological systems. Annu. Rev. Biomed. Eng. 2017;19:249–277. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
