

Comparative Development of the Ant Chemosensory System
- PMID: 32559450
- PMCID: PMC7438299
- DOI: 10.1016/j.cub.2020.05.072
Comparative Development of the Ant Chemosensory System
Abstract
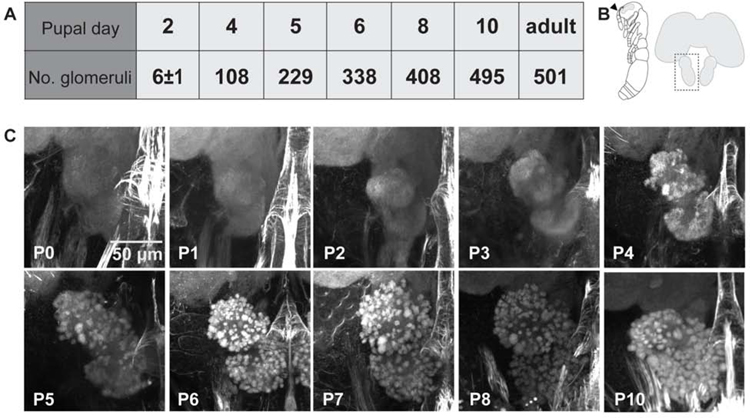
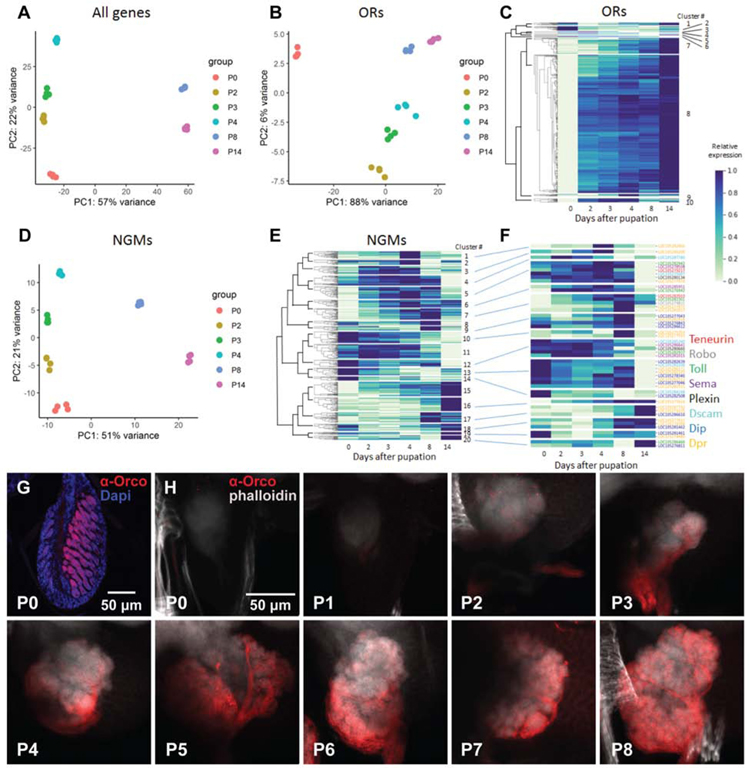
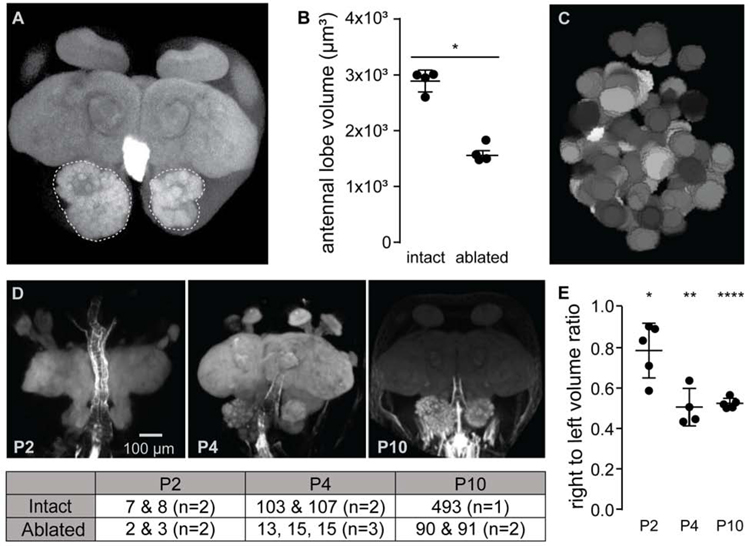
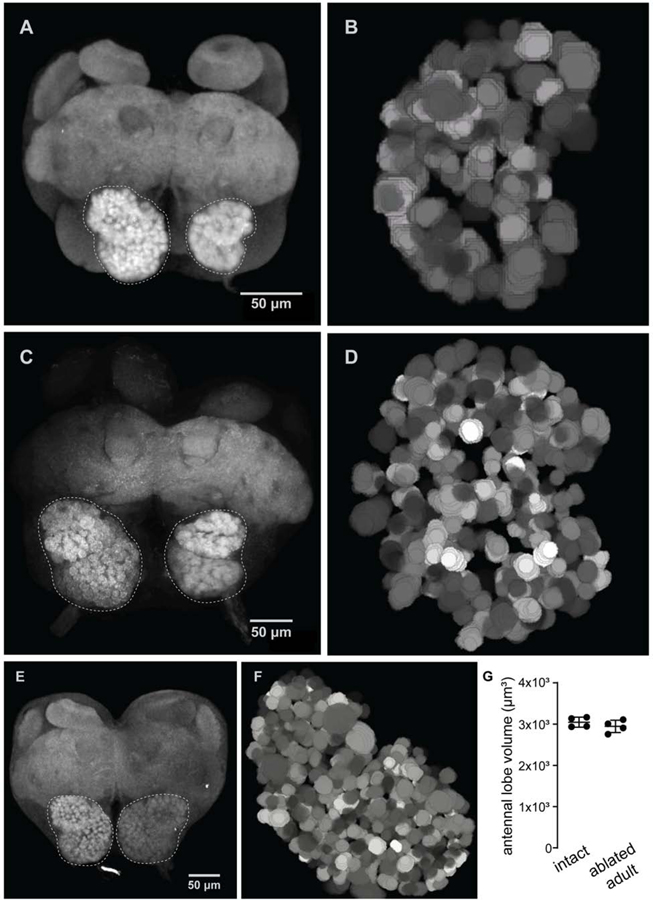
The insect antennal lobe (AL) contains the first synapses of the olfactory system, where olfactory sensory neurons (OSNs) contact second-order projection neurons (PNs). In Drosophila melanogaster, OSNs expressing specific receptor genes send stereotyped projections to one or two of about 50 morphologically defined glomeruli [1-3]. The mechanisms for this precise matching between OSNs and PNs have been studied extensively in D. melanogaster, where development is deterministic and independent of neural activity [4-6]. However, a number of insect lineages, most notably the ants, have receptor gene repertoires many times larger than D. melanogaster and exhibit more structurally complex antennal lobes [7-12]. Moreover, perturbation of OSN function via knockout of the odorant receptor (OR) co-receptor, Orco, results in drastic AL reductions in ants [13, 14], but not in Drosophila [15]. Here, we characterize AL development in the clonal raider ant, Ooceraea biroi. We find that, unlike in Drosophila, ORs and Orco are expressed before the onset of glomerulus formation, and Orco protein is trafficked to developing axon terminals, raising the possibility that ORs play a role during ant AL development. Additionally, ablating ant antennae at the onset of pupation results in AL defects that recapitulate the Orco mutant phenotype. Thus, early loss of functional OSN innervation reveals latent structure in the AL that develops independently of peripheral input, suggesting that the AL is initially pre-patterned and later refined in an OSN-dependent manner. This two-step process might increase developmental flexibility and thereby facilitate the rapid evolution and expansion of the ant chemosensory system.
Keywords: Formicidae; Ooceraea biroi; antennal lobe; chemosensation; clonal raider ant; odorant receptors; olfaction; pheromones; sensory systems; social insects.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Comment in
-
Ant Olfaction: Smells Like an Insect, Develops Like a Mammal.Curr Biol. 2020 Aug 17;30(16):R950-R952. doi: 10.1016/j.cub.2020.06.074. Curr Biol. 2020. PMID: 32810458
References
-
- Laissue PP, Reiter C, Hiesinger PR, Halter S, Fischbach KF, and Stocker RF (1999). Three-dimensional reconstruction of the antennal lobe in Drosophila melanogaster. Journal of Comparative Neurology 405, 543–552. - PubMed
-
- Couto A, Alenius M, and Dickson BJ (2005). Molecular, Anatomical, and Functional Organization of the Drosophila Olfactory System. Current Biology 15, 1535–1547. - PubMed
-
- Vosshall LB (2000). Olfaction in Drosophila. Current Opinion in Neurobiology 10, 498–503. - PubMed
-
- Sardana J, Organisti C, and Kadow ICG (2018). Eph Receptor Effector Ephexin Mediates Olfactory Dendrite Targeting in Drosophila. Developmental Neurobiology 78, 873–888. - PubMed
-
- Technau GM ed. (2008). Brain development in Drosophila melanogaster (New York : Austin, Tex: Springer Science+Business Media ; Landes Bioscience; ).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
