

Mutually stabilizing interactions between proto-peptides and RNA
- PMID: 32561731
- PMCID: PMC7305224
- DOI: 10.1038/s41467-020-16891-5
Mutually stabilizing interactions between proto-peptides and RNA
Abstract
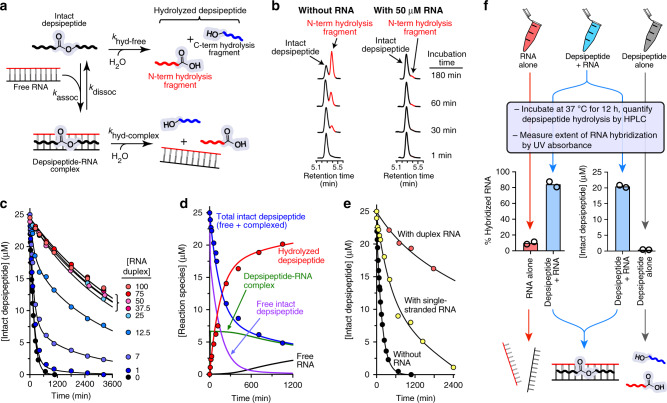
The close synergy between peptides and nucleic acids in current biology is suggestive of a functional co-evolution between the two polymers. Here we show that cationic proto-peptides (depsipeptides and polyesters), either produced as mixtures from plausibly prebiotic dry-down reactions or synthetically prepared in pure form, can engage in direct interactions with RNA resulting in mutual stabilization. Cationic proto-peptides significantly increase the thermal stability of folded RNA structures. In turn, RNA increases the lifetime of a depsipeptide by >30-fold. Proto-peptides containing the proteinaceous amino acids Lys, Arg, or His adjacent to backbone ester bonds generally promote RNA duplex thermal stability to a greater magnitude than do analogous sequences containing non-proteinaceous residues. Our findings support a model in which tightly-intertwined biological dependencies of RNA and protein reflect a long co-evolutionary history that began with rudimentary, mutually-stabilizing interactions at early stages of polypeptide and nucleic acid co-existence.
Conflict of interest statement
The authors declare no competing interests.
Figures
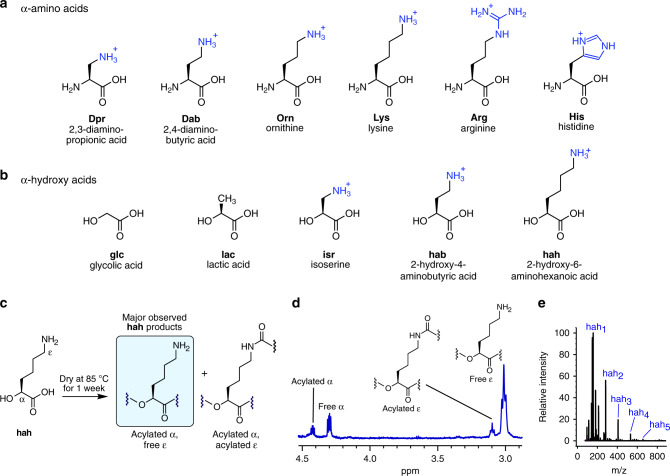
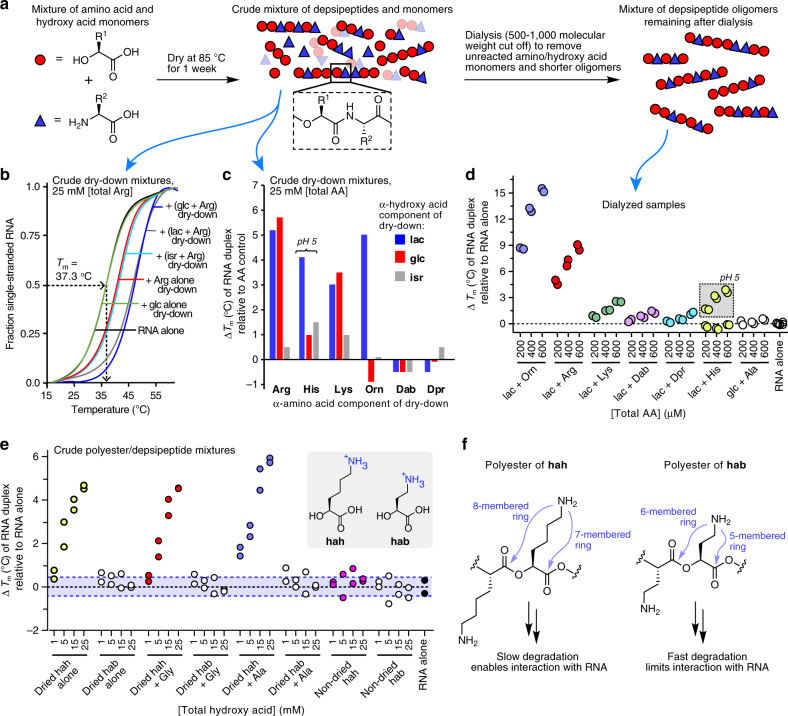
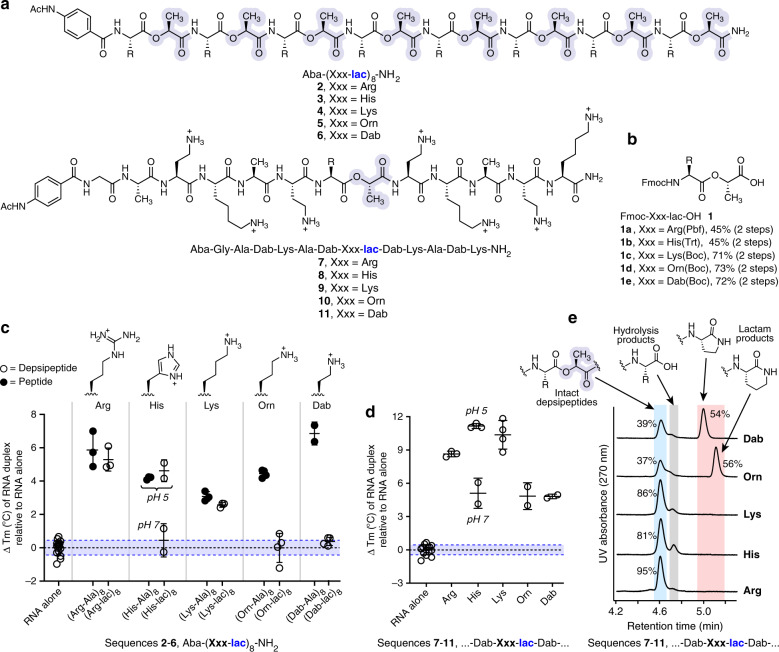
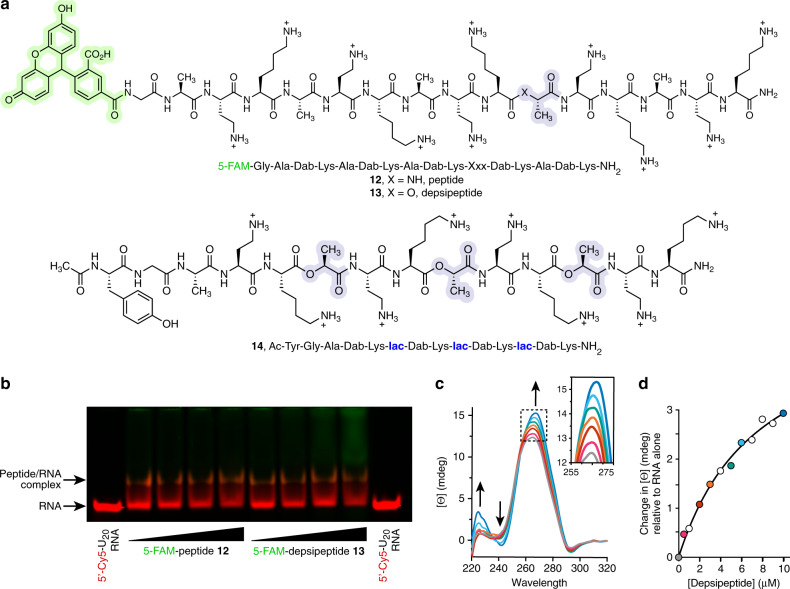
References
-
- Frenkel-Pinter M, Samanta M, Ashkenasy G, Leman LJ. Prebiotic peptides: Molecular hubs in the origin of life. Chem. Rev. 2020;120:4707–4765. - PubMed
-
- Orgel LE. Prebiotic chemistry and the origin of the RNA world. Crit. Rev. Biochem. Mol. Biol. 2004;39:99–123. - PubMed
-
- Neveu M, Kim HJ, Benner SA. The “strong” RNA world hypothesis: fifty years old. Astrobiology. 2013;13:391–403. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
