

The bacterial copper resistance protein CopG contains a cysteine-bridged tetranuclear copper cluster
- PMID: 32571874
- PMCID: PMC7415965
- DOI: 10.1074/jbc.RA120.013907
The bacterial copper resistance protein CopG contains a cysteine-bridged tetranuclear copper cluster
Abstract
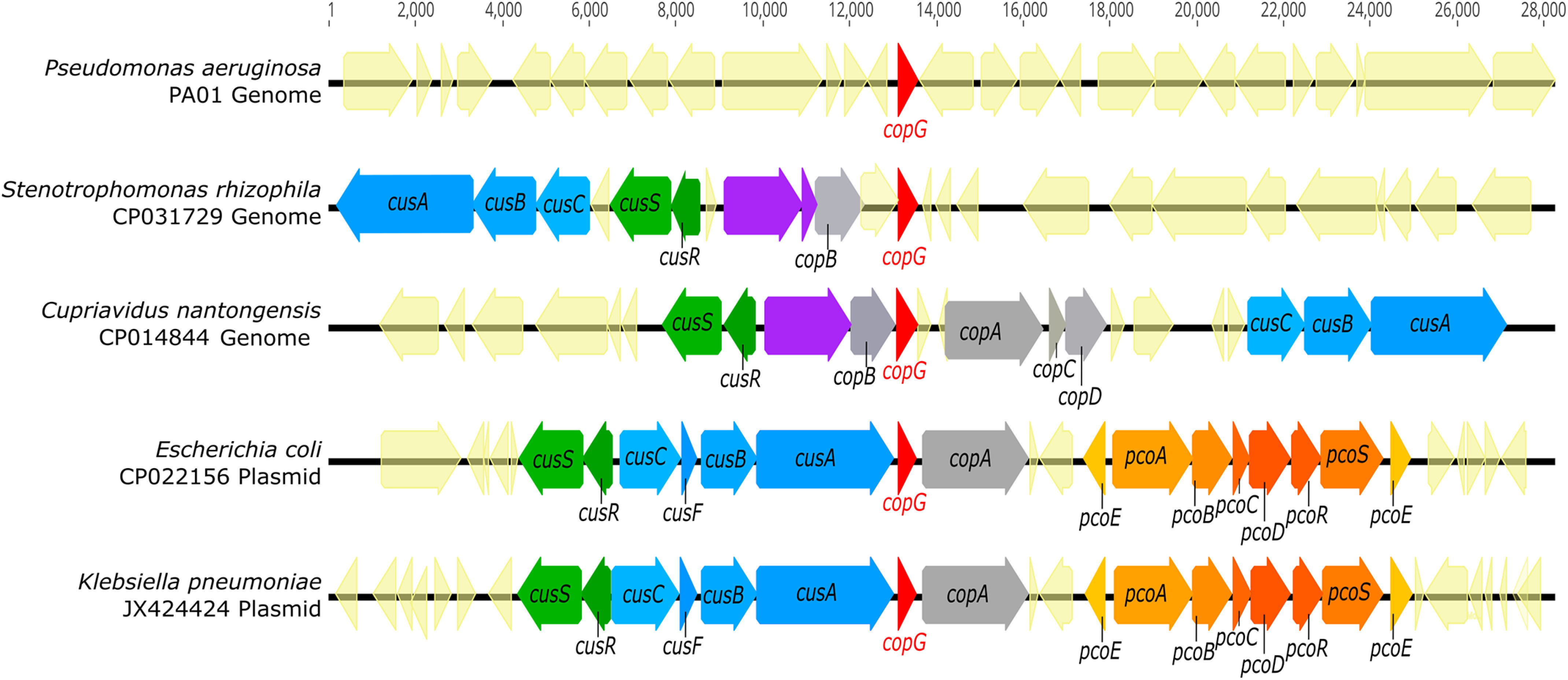
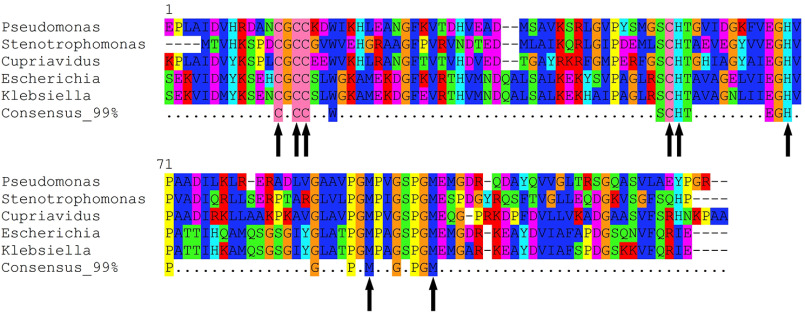
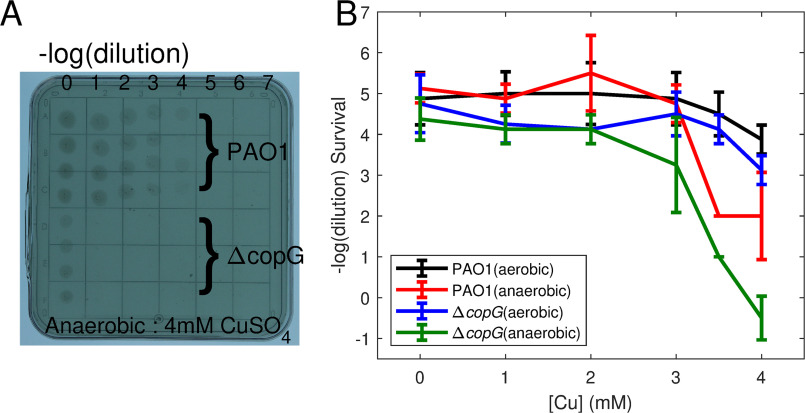
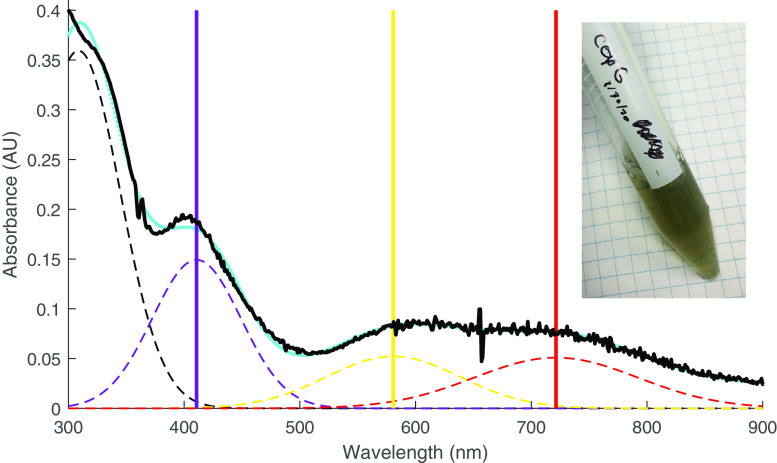
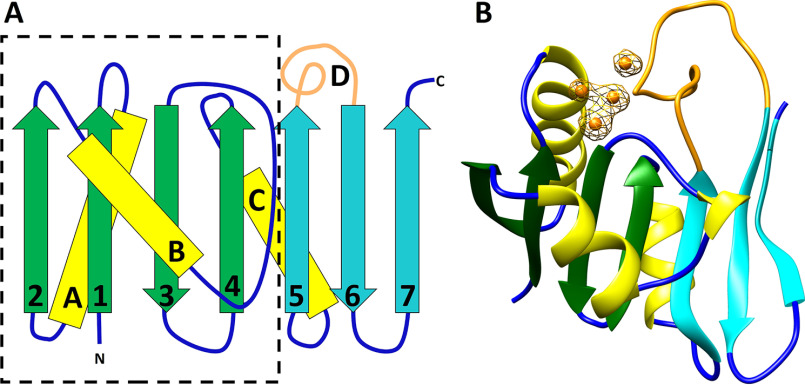
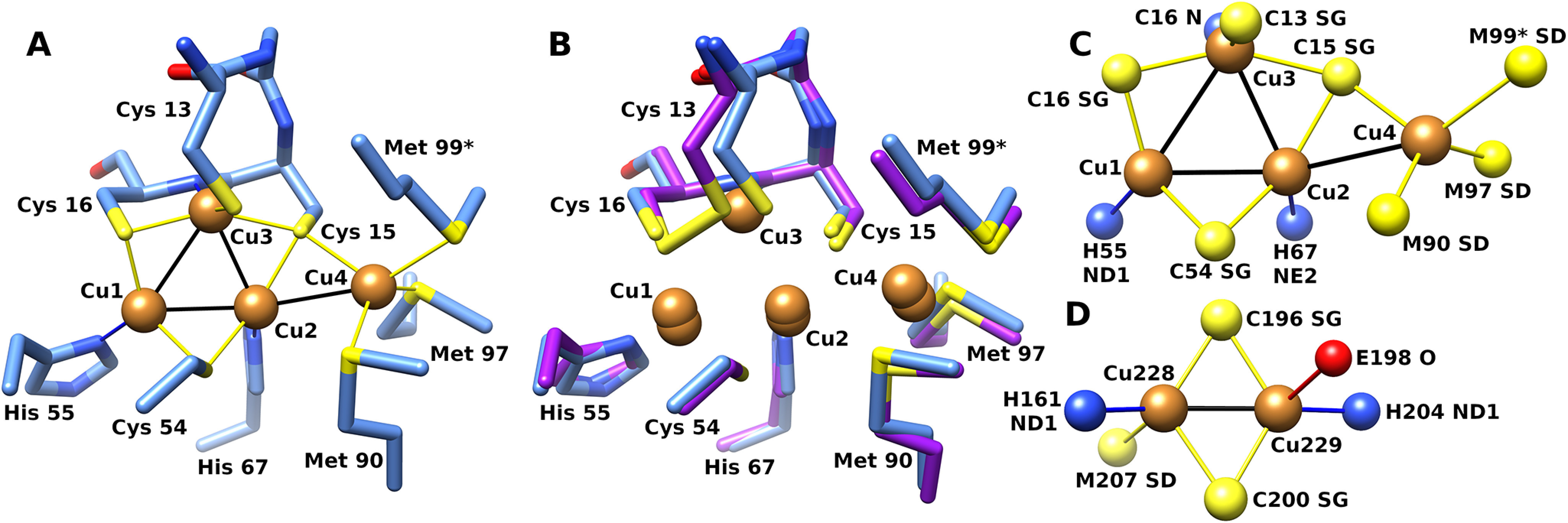
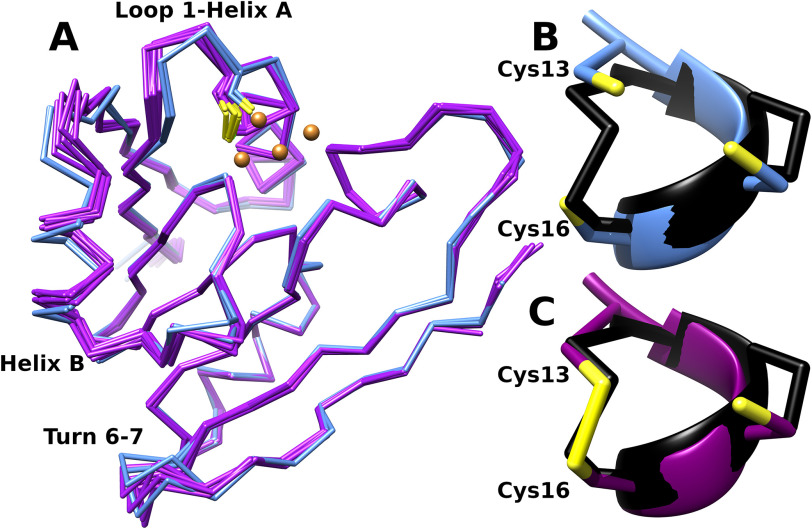
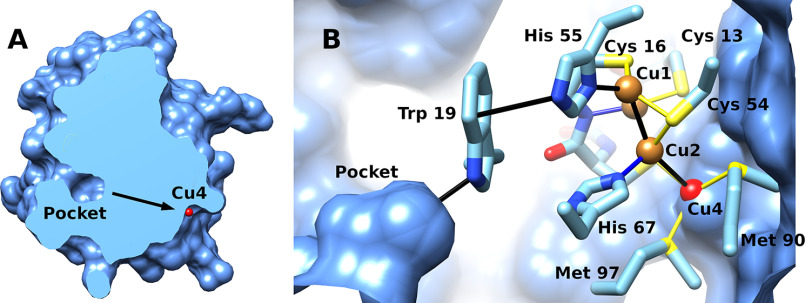
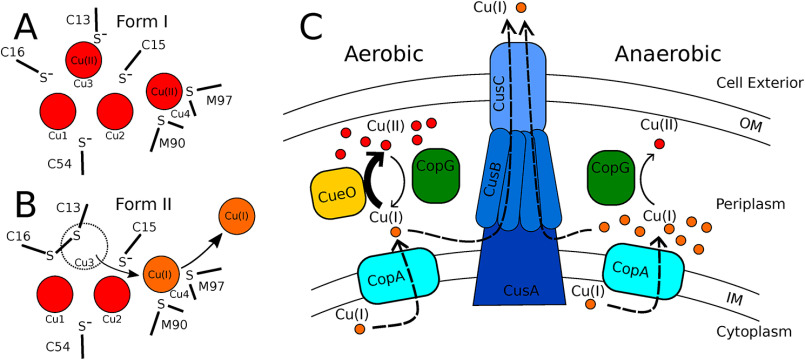
CopG is an uncharacterized protein ubiquitous in Gram-negative bacteria whose gene frequently occurs in clusters of copper resistance genes and can be recognized by the presence of a conserved CxCC motif. To investigate its contribution to copper resistance, here we undertook a structural and biochemical characterization of the CopG protein from Pseudomonas aeruginosa Results from biochemical analyses of CopG purified under aerobic conditions indicate that it is a green copper-binding protein that displays absorbance maxima near 411, 581, and 721 nm and is monomeric in solution. Determination of the three-dimensional structure by X-ray crystallography revealed that CopG consists of a thioredoxin domain with a C-terminal extension that contributes to metal binding. We noted that adjacent to the CxCC motif is a cluster of four copper ions bridged by cysteine sulfur atoms. Structures of CopG in two oxidation states support the assignment of this protein as an oxidoreductase. On the basis of these structural and spectroscopic findings and also genetic evidence, we propose that CopG has a role in interconverting Cu(I) and Cu(II) to minimize toxic effects and facilitate export by the Cus RND transporter efflux system.
Keywords: P. aeruginosa; Pseudomonas aeruginosa; copper; metal homeostasis; metal ion homeostasis; metal resistance; metalloprotein; oxidation-reduction; oxidation-reduction (redox); structure-function; thioredoxin.
© 2020 Hausrath et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Lazary A., Weinberg I., Vatine J. J., Jefidoff A., Bardenstein R., Borkow G., and Ohana N. (2014) Reduction of healthcare-associated infections in a long-term care brain injury ward by replacing regular linens with biocidal copper oxide impregnated linens. Int. J. Infect. Dis. 24, 23–29 10.1016/j.ijid.2014.01.022 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
