

Structural and functional conservation of the programmed -1 ribosomal frameshift signal of SARS coronavirus 2 (SARS-CoV-2)
- PMID: 32571880
- PMCID: PMC7397099
- DOI: 10.1074/jbc.AC120.013449
Structural and functional conservation of the programmed -1 ribosomal frameshift signal of SARS coronavirus 2 (SARS-CoV-2)
Abstract
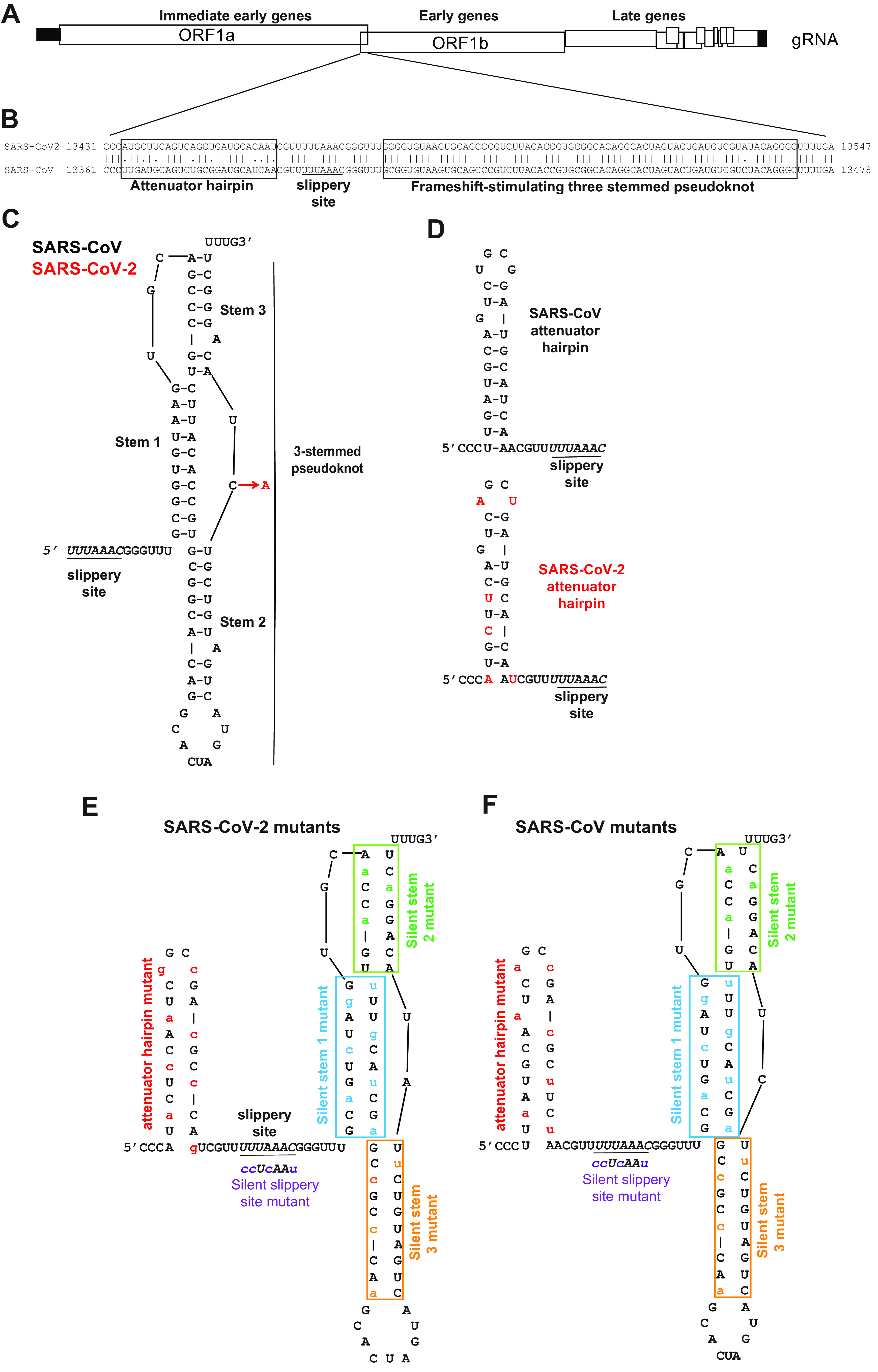
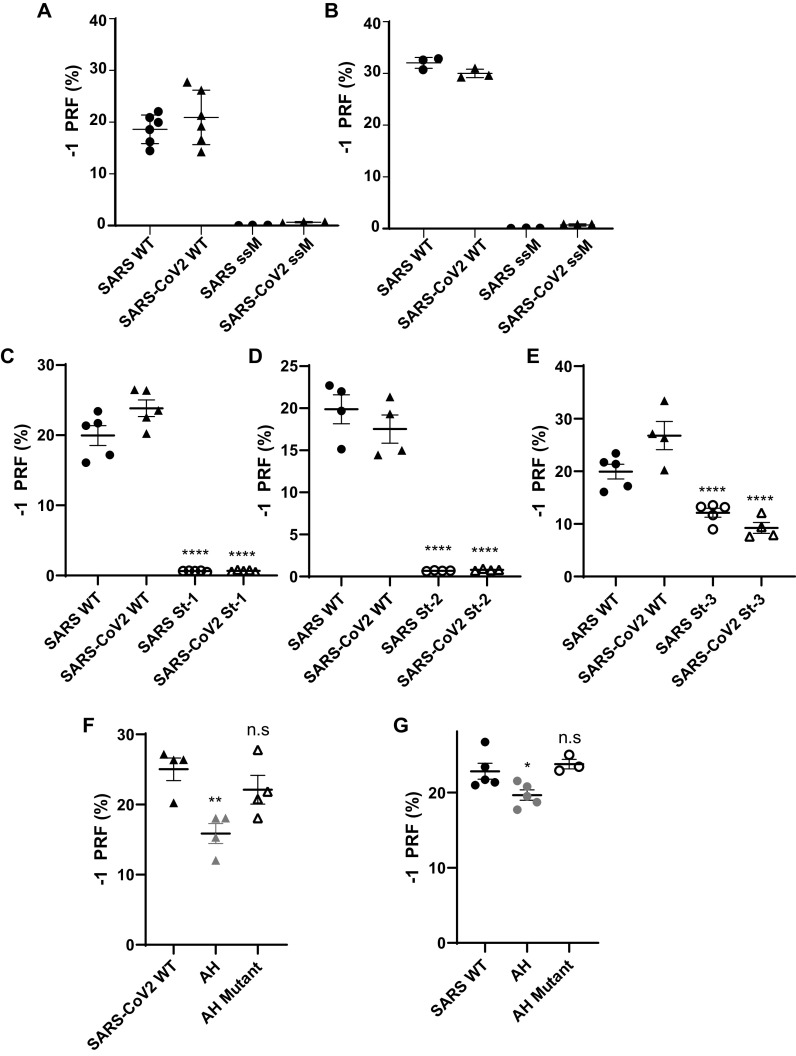
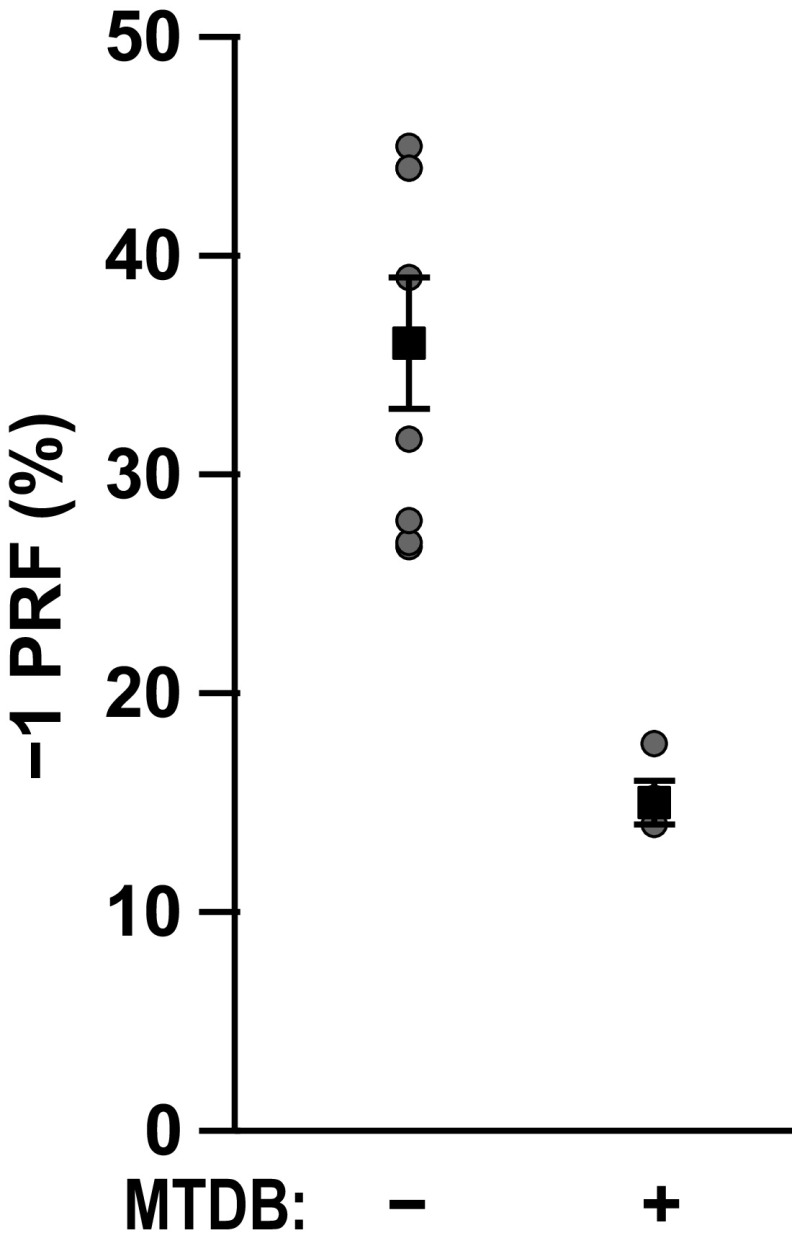
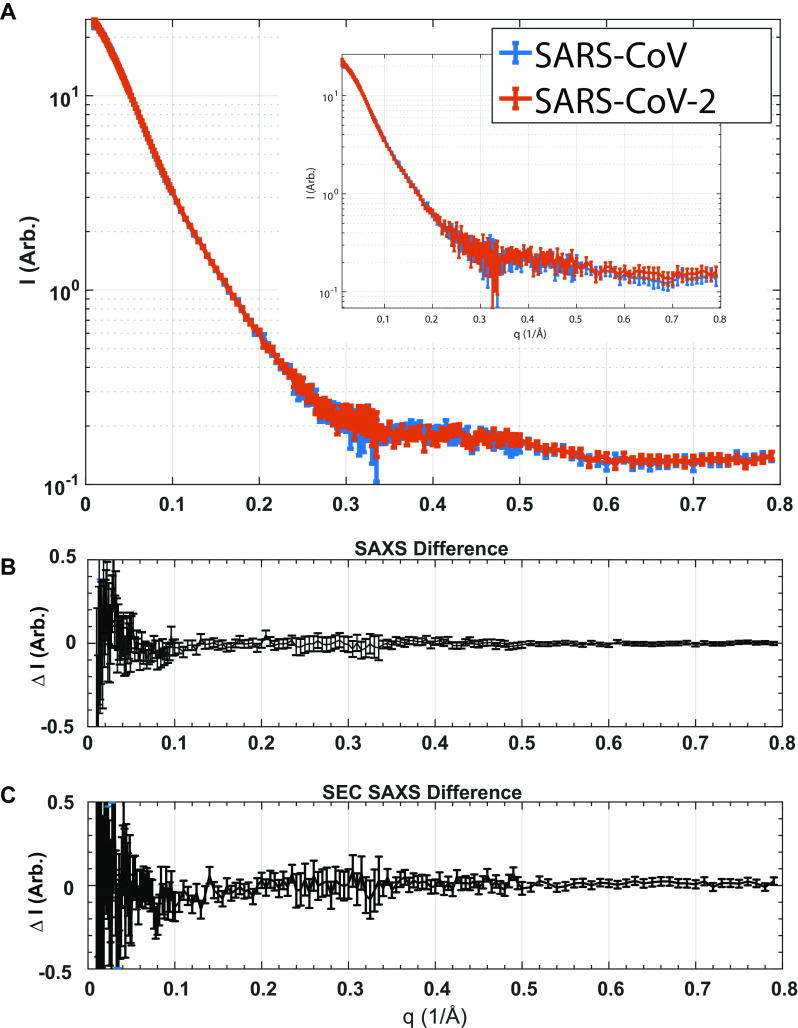
Approximately 17 years after the severe acute respiratory syndrome coronavirus (SARS-CoV) epidemic, the world is currently facing the COVID-19 pandemic caused by SARS corona virus 2 (SARS-CoV-2). According to the most optimistic projections, it will take more than a year to develop a vaccine, so the best short-term strategy may lie in identifying virus-specific targets for small molecule-based interventions. All coronaviruses utilize a molecular mechanism called programmed -1 ribosomal frameshift (-1 PRF) to control the relative expression of their proteins. Previous analyses of SARS-CoV have revealed that it employs a structurally unique three-stemmed mRNA pseudoknot that stimulates high -1 PRF rates and that it also harbors a -1 PRF attenuation element. Altering -1 PRF activity impairs virus replication, suggesting that this activity may be therapeutically targeted. Here, we comparatively analyzed the SARS-CoV and SARS-CoV-2 frameshift signals. Structural and functional analyses revealed that both elements promote similar -1 PRF rates and that silent coding mutations in the slippery sites and in all three stems of the pseudoknot strongly ablate -1 PRF activity. We noted that the upstream attenuator hairpin activity is also functionally retained in both viruses, despite differences in the primary sequence in this region. Small-angle X-ray scattering analyses indicated that the pseudoknots in SARS-CoV and SARS-CoV-2 have the same conformation. Finally, a small molecule previously shown to bind the SARS-CoV pseudoknot and inhibit -1 PRF was similarly effective against -1 PRF in SARS-CoV-2, suggesting that such frameshift inhibitors may be promising lead compounds to combat the current COVID-19 pandemic.
Keywords: (+) ssRNA; RNA; RNA structure; coronavirus; coronavirus disease 2019 (COVID-19); inhibitor; mRNA pseudoknot; programmed −1 ribosomal frameshifting (−1 PRF); small molecule inhibitor; translation; virus.
© 2020 Kelly et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Update of
-
Structural and functional conservation of the programmed -1 ribosomal frameshift signal of SARS-CoV-2.bioRxiv [Preprint]. 2020 Jun 15:2020.03.13.991083. doi: 10.1101/2020.03.13.991083. bioRxiv. 2020. Update in: J Biol Chem. 2020 Jul 31;295(31):10741-10748. doi: 10.1074/jbc.AC120.013449. PMID: 32587971 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
