

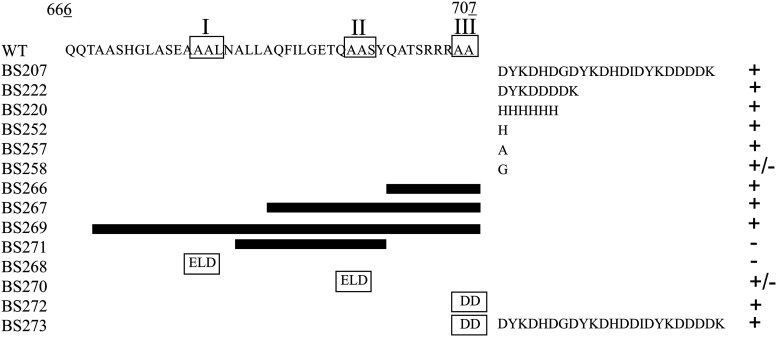
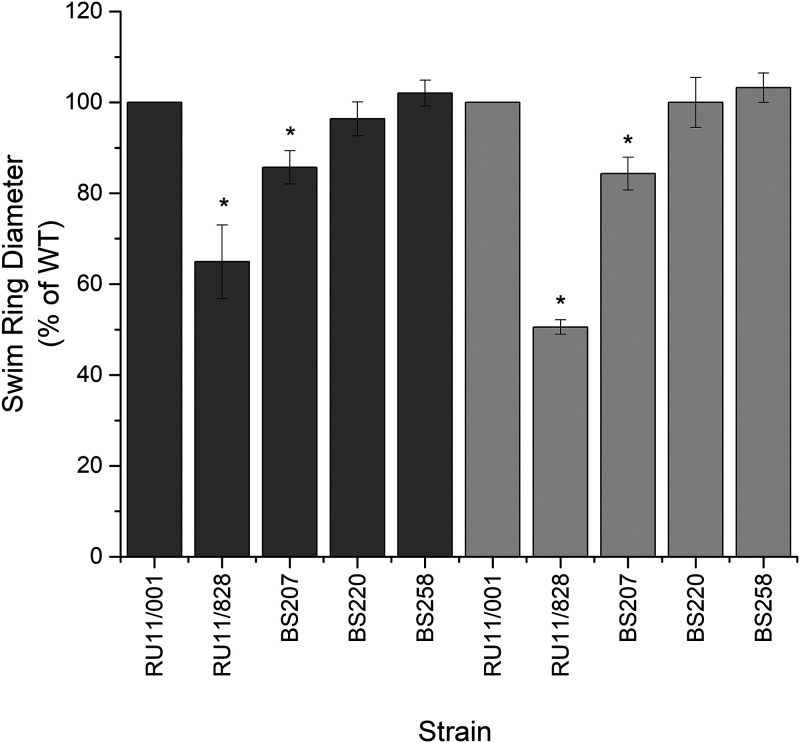
Programmed Proteolysis of Chemotaxis Proteins in Sinorhizobium meliloti: Features in the C-Terminal Region Control McpU Degradation
- PMID: 32571966
- PMCID: PMC7417835
- DOI: 10.1128/JB.00124-20
Programmed Proteolysis of Chemotaxis Proteins in Sinorhizobium meliloti: Features in the C-Terminal Region Control McpU Degradation
Abstract
Chemotaxis and motility are important traits that support bacterial survival in various ecological niches and in pathogenic and symbiotic host interaction. Chemotactic stimuli are sensed by chemoreceptors or
Keywords: ClpXP protease; alphaproteobacteria; cell cycle; chemotaxis; epitope tags.
Copyright © 2020 American Society for Microbiology.
Figures





Similar articles
-
Cellular Stoichiometry of Methyl-Accepting Chemotaxis Proteins in Sinorhizobium meliloti.J Bacteriol. 2018 Feb 23;200(6):e00614-17. doi: 10.1128/JB.00614-17. Print 2018 Mar 15. J Bacteriol. 2018. PMID: 29263102 Free PMC article.
-
Sinorhizobium meliloti chemoreceptor McpU mediates chemotaxis toward host plant exudates through direct proline sensing.Appl Environ Microbiol. 2014 Jun;80(11):3404-15. doi: 10.1128/AEM.00115-14. Epub 2014 Mar 21. Appl Environ Microbiol. 2014. PMID: 24657863 Free PMC article.
-
McpT, a Broad-Range Carboxylate Chemoreceptor in Sinorhizobium meliloti.J Bacteriol. 2021 Aug 9;203(17):e0021621. doi: 10.1128/JB.00216-21. Epub 2021 Aug 9. J Bacteriol. 2021. PMID: 34124939 Free PMC article.
-
Coordination of symbiosis and cell cycle functions in Sinorhizobium meliloti.Biochim Biophys Acta Gene Regul Mech. 2019 Jul;1862(7):691-696. doi: 10.1016/j.bbagrm.2018.05.003. Epub 2018 May 19. Biochim Biophys Acta Gene Regul Mech. 2019. PMID: 29783033 Review.
-
Sensory transduction to the flagellar motor of Sinorhizobium meliloti.J Mol Microbiol Biotechnol. 2002 May;4(3):183-6. J Mol Microbiol Biotechnol. 2002. PMID: 11931544 Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
