

The Tomato DELLA Protein PROCERA Promotes Abscisic Acid Responses in Guard Cells by Upregulating an Abscisic Acid Transporter
- PMID: 32576645
- PMCID: PMC7479916
- DOI: 10.1104/pp.20.00485
The Tomato DELLA Protein PROCERA Promotes Abscisic Acid Responses in Guard Cells by Upregulating an Abscisic Acid Transporter
Abstract
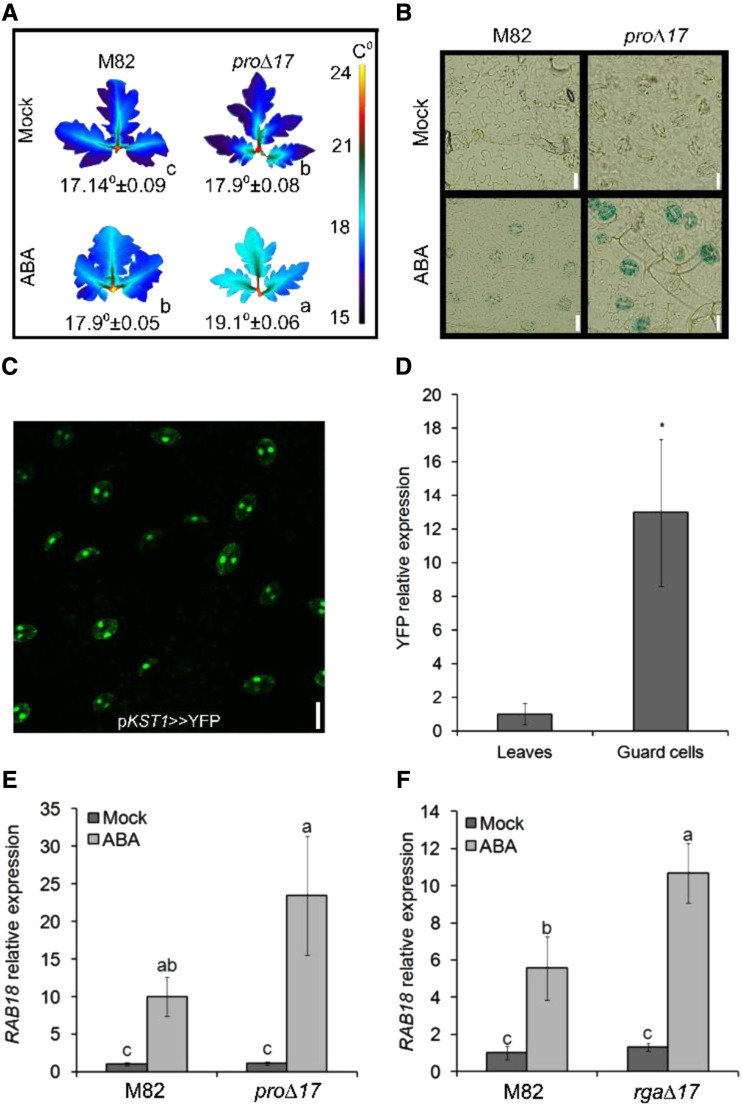
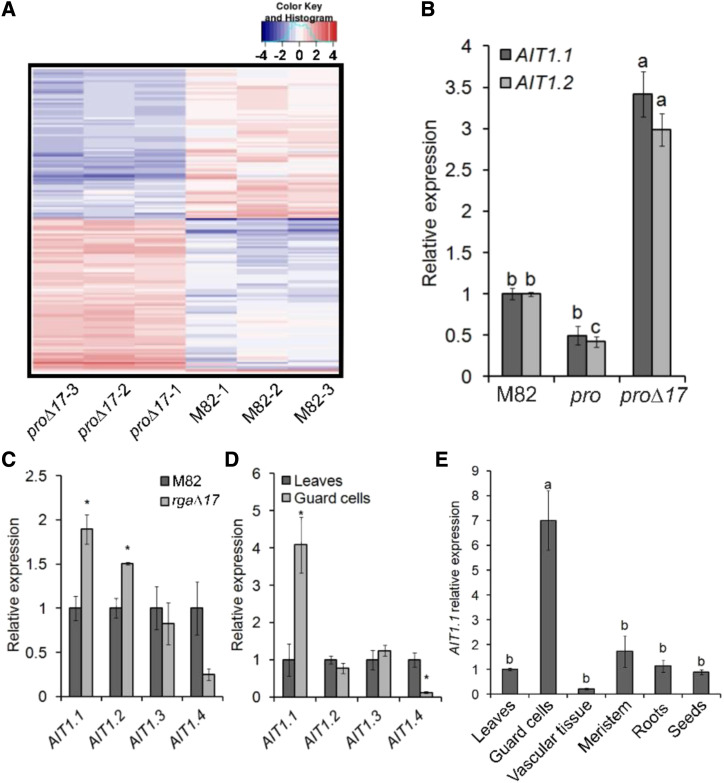
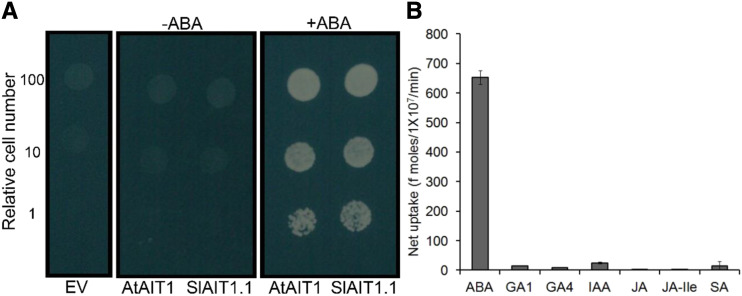
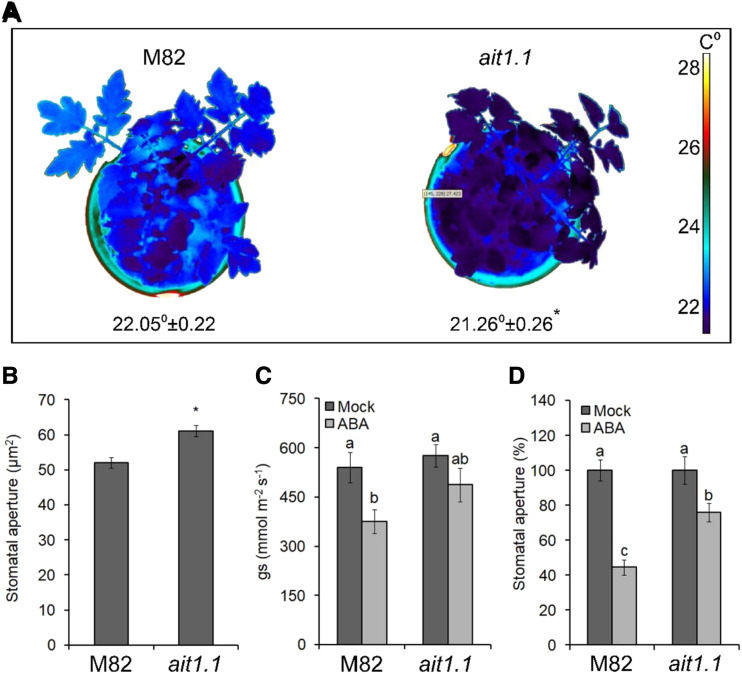
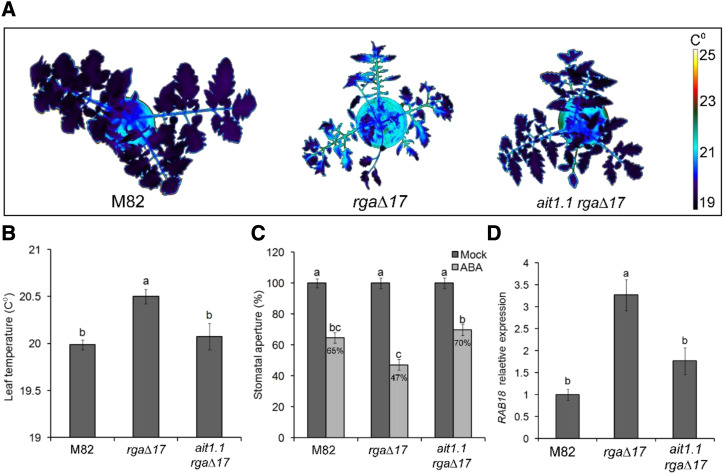
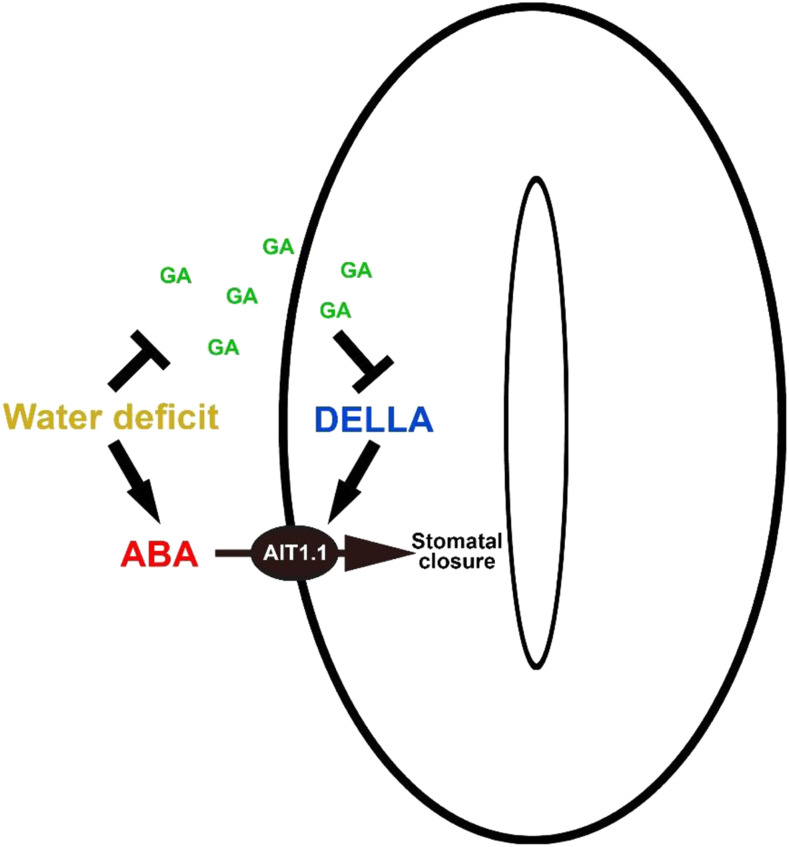
Plants reduce transpiration through stomatal closure to avoid drought stress. While abscisic acid (ABA) has a central role in the regulation of stomatal closure under water-deficit conditions, we demonstrated in tomato (Solanum lycopersicum) that a gibberellin response inhibitor, the DELLA protein PROCERA (PRO), promotes ABA-induced stomatal closure and gene transcription in guard cells. To study how PRO affects stomatal closure, we performed RNA-sequencing analysis of isolated guard cells and identified the ABA transporters ABA-IMPORTING TRANSPORTER1 1 (AIT1 1) and AIT1 2, also called NITRATE TRANSPORTER1/PTR TRANSPORTER FAMILY4 6 in Arabidopsis (Arabidopsis thaliana), as being upregulated by PRO. Tomato has four AIT1 genes, but only AIT1 1 and AIT1 2 were upregulated by PRO, and only AIT1 1 exhibited high expression in guard cells. Functional analysis of AIT1 1 in yeast (Saccharomyces cerevisiae) confirmed its activity as an ABA transporter, possibly an importer. A clustered regularly interspaced short palindromic repeats-Cas9-derived ait1 1 mutant exhibited an increased transpiration, a larger stomatal aperture, and a reduced stomatal response to ABA. Moreover, ait1 1 suppressed the promoting effects of PRO on ABA-induced stomatal closure and gene expression in guard cells, suggesting that the effects of PRO on stomatal aperture and transpiration are AIT1.1-dependent. Previous studies suggest a negative crosstalk between gibberellin and ABA that is mediated by changes in hormone biosynthesis and signaling. The results of this study suggest this crosstalk is also mediated by changes in hormone transport.
© 2020 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, van der Straeten D, Peng J, Harberd NP(2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311: 91–94 - PubMed
-
- Bassel GW, Mullen RT, Bewley JD(2008) Procera is a putative DELLA mutant in tomato (Solanum lycopersicum): Effects on the seed and vegetative plant. J Exp Bot 59: 585–593 - PubMed
-
- Bauer H, Ache P, Lautner S, Fromm J, Hartung W, Al-Rasheid KA, Sonnewald S, Sonnewald U, Kneitz S, Lachmann N, et al. (2013) The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr Biol 23: 53–57 - PubMed
-
- Benjamini Y, Hochberg Y(1995) Controlling the false discovery rate: A practical and powerful approach to multiple testing. J R Stat Soc 57: 289–300
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
