

Multifaceted deregulation of gene expression and protein synthesis with age
- PMID: 32576685
- PMCID: PMC7354943
- DOI: 10.1073/pnas.2001788117
Multifaceted deregulation of gene expression and protein synthesis with age
Abstract
Protein synthesis represents a major metabolic activity of the cell. However, how it is affected by aging and how this in turn impacts cell function remains largely unexplored. To address this question, herein we characterized age-related changes in both the transcriptome and translatome of mouse tissues over the entire life span. We showed that the transcriptome changes govern those in the translatome and are associated with altered expression of genes involved in inflammation, extracellular matrix, and lipid metabolism. We also identified genes that may serve as candidate biomarkers of aging. At the translational level, we uncovered sustained down-regulation of a set of 5'-terminal oligopyrimidine (5'-TOP) transcripts encoding protein synthesis and ribosome biogenesis machinery and regulated by the mTOR pathway. For many of them, ribosome occupancy dropped twofold or even more. Moreover, with age, ribosome coverage gradually decreased in the vicinity of start codons and increased near stop codons, revealing complex age-related changes in the translation process. Taken together, our results reveal systematic and multidimensional deregulation of protein synthesis, showing how this major cellular process declines with age.
Keywords: 5′-TOP; aging; proteostasis; ribosome profiling; transcriptome.
Conflict of interest statement
The authors declare no competing interest.
Figures
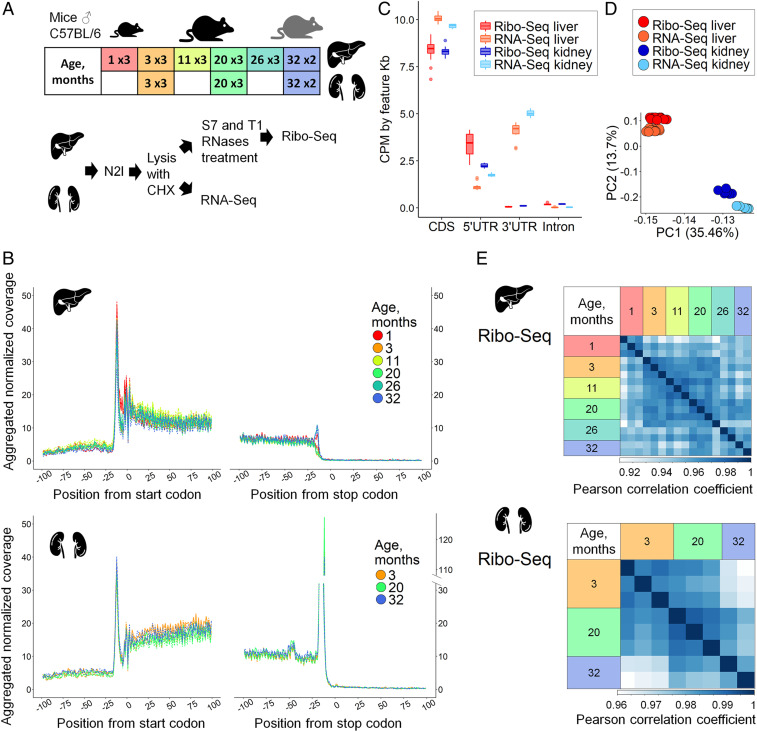
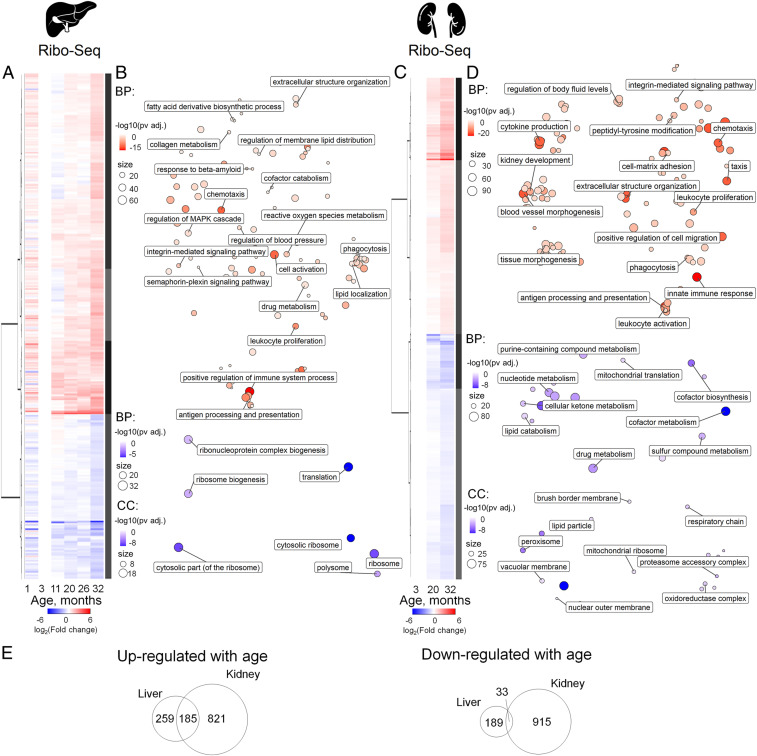
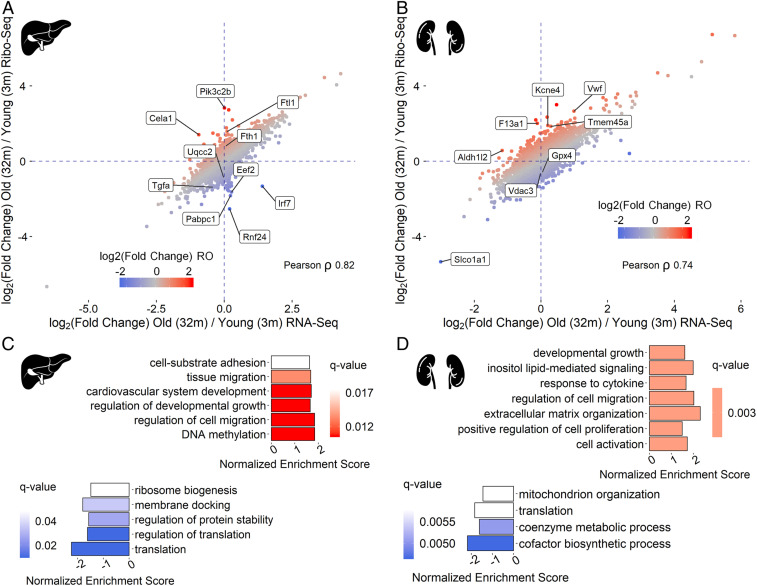
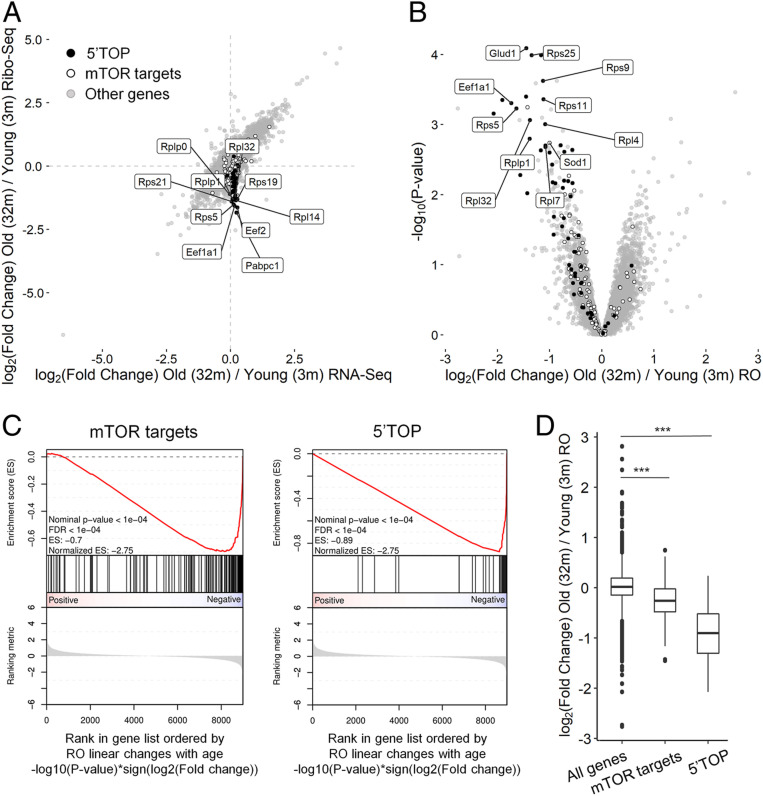
References
-
- Hipp M. S., Kasturi P., Hartl F. U., The proteostasis network and its decline in ageing. Nat. Rev. Mol. Cell Biol. 20, 421–435 (2019). - PubMed
-
- Steffen K. K., Dillin A., A ribosomal perspective on proteostasis and aging. Cell Metab. 23, 1004–1012 (2016). - PubMed
-
- Gerashchenko M. V., Peterfi Z., Gladyshev V. N., Organ-specific translation elongation rates measured by in vivo ribosome profiling. bioRxiv:279257 (8 March 2018).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
