

The clonal evolution of metastatic colorectal cancer
- PMID: 32577507
- PMCID: PMC7286679
- DOI: 10.1126/sciadv.aay9691
The clonal evolution of metastatic colorectal cancer
Abstract
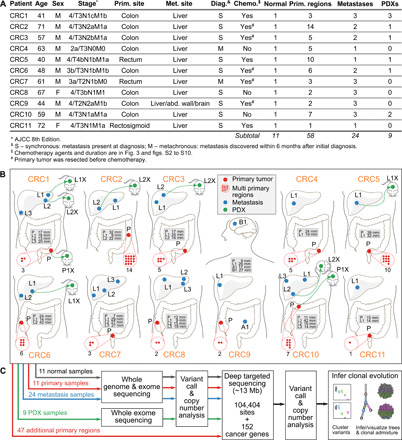
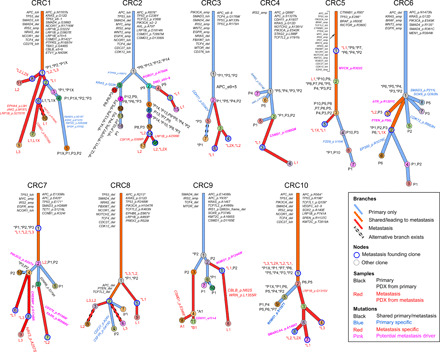
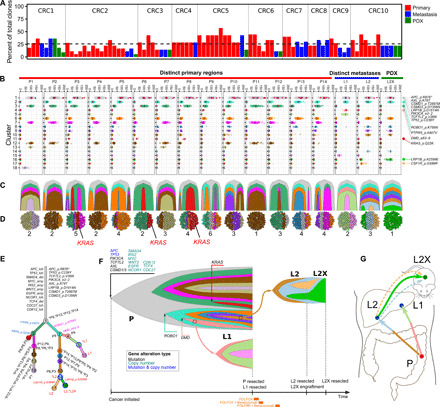
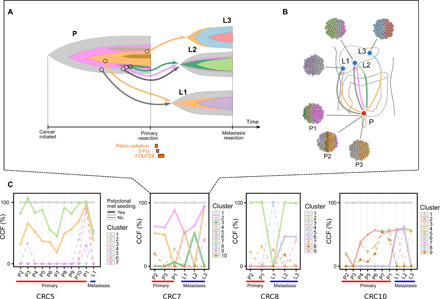
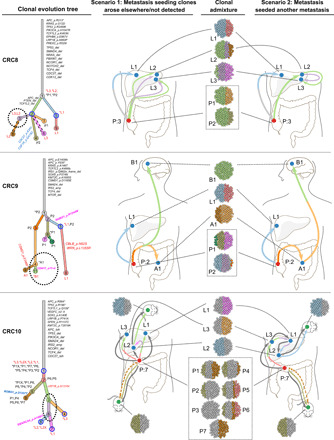
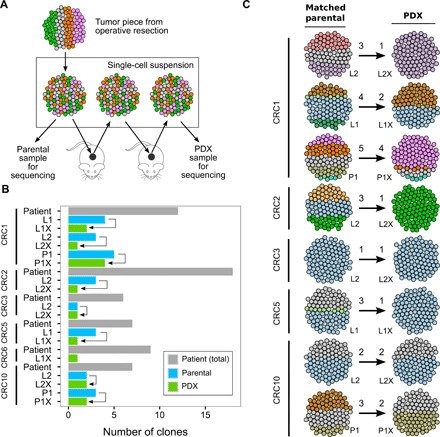
Tumor heterogeneity and evolution drive treatment resistance in metastatic colorectal cancer (mCRC). Patient-derived xenografts (PDXs) can model mCRC biology; however, their ability to accurately mimic human tumor heterogeneity is unclear. Current genomic studies in mCRC have limited scope and lack matched PDXs. Therefore, the landscape of tumor heterogeneity and its impact on the evolution of metastasis and PDXs remain undefined. We performed whole-genome, deep exome, and targeted validation sequencing of multiple primary regions, matched distant metastases, and PDXs from 11 patients with mCRC. We observed intricate clonal heterogeneity and evolution affecting metastasis dissemination and PDX clonal selection. Metastasis formation followed both monoclonal and polyclonal seeding models. In four cases, metastasis-seeding clones were not identified in any primary region, consistent with a metastasis-seeding-metastasis model. PDXs underrepresented the subclonal heterogeneity of parental tumors. These suggest that single sample tumor sequencing and current PDX models may be insufficient to guide precision medicine.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Fearon E. R., Vogelstein B., A genetic model for colorectal tumorigenesis. Cell 61, 759–767 (1990). - PubMed
-
- Brannon A. R., Vakiani E., Sylvester B. E., Scott S. N., McDermott G., Shah R. H., Kania K., Viale A., Oschwald D. M., Vacic V., Emde A.-K., Cercek A., Yaeger R., Kemeny N. E., Saltz L. B., Shia J., D’Angelica M. I., Weiser M. R., Solit D. B., Berger M. F., Comparative sequencing analysis reveals high genomic concordance between matched primary and metastatic colorectal cancer lesions. Genome Biol. 15, 454 (2014). - PMC - PubMed
-
- Wei Q., Ye Z., Zhong X., Li L., Wang C., Myers R. E., Palazzo J. P., Fortuna D., Yan A., Waldman S. A., Chen X., Posey J. A., Basu-Mallick A., Jiang B. H., Hou L., Shu J., Sun Y., Xing J., Li B., Yang H., Multiregion whole-exome sequencing of matched primary and metastatic tumors revealed genomic heterogeneity and suggested polyclonal seeding in colorectal cancer metastasis. Ann. Oncol. 28, 2135–2141 (2017). - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
