

A polybasic domain in aPKC mediates Par6-dependent control of membrane targeting and kinase activity
- PMID: 32580209
- PMCID: PMC7337507
- DOI: 10.1083/jcb.201903031
A polybasic domain in aPKC mediates Par6-dependent control of membrane targeting and kinase activity
Abstract
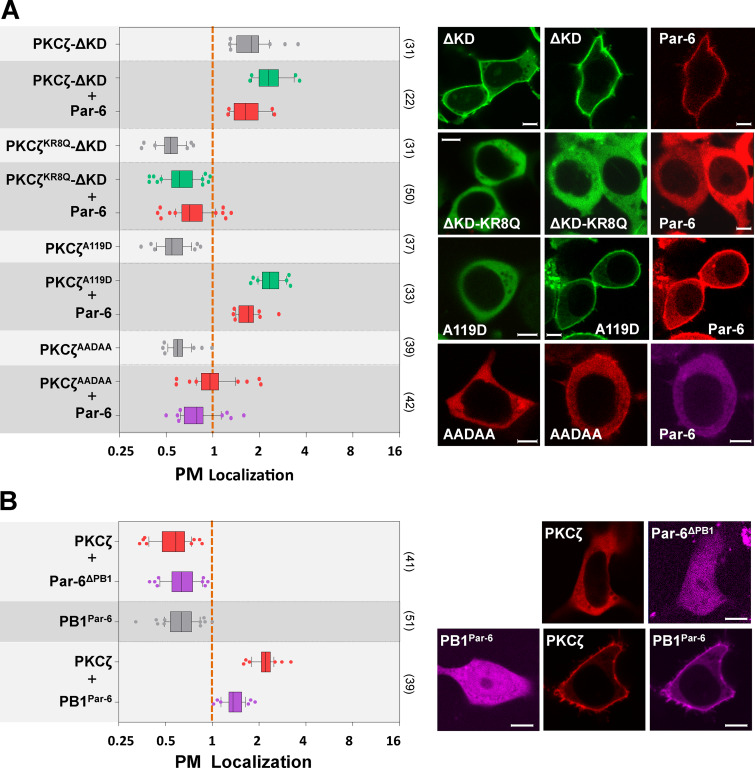
Mechanisms coupling the atypical PKC (aPKC) kinase activity to its subcellular localization are essential for cell polarization. Unlike other members of the PKC family, aPKC has no well-defined plasma membrane (PM) or calcium binding domains, leading to the assumption that its subcellular localization relies exclusively on protein-protein interactions. Here we show that in both Drosophila and mammalian cells, the pseudosubstrate region (PSr) of aPKC acts as a polybasic domain capable of targeting aPKC to the PM via electrostatic binding to PM PI4P and PI(4,5)P2. However, physical interaction between aPKC and Par-6 is required for the PM-targeting of aPKC, likely by allosterically exposing the PSr to bind PM. Binding of Par-6 also inhibits aPKC kinase activity, and such inhibition can be relieved through Par-6 interaction with apical polarity protein Crumbs. Our data suggest a potential mechanism in which allosteric regulation of polybasic PSr by Par-6 couples the control of both aPKC subcellular localization and spatial activation of its kinase activity.
© 2020 Dong et al.
Figures












Similar articles
-
A PDZ-kinase allosteric relay mediates Par complex regulator exchange.J Biol Chem. 2025 Feb;301(2):108097. doi: 10.1016/j.jbc.2024.108097. Epub 2024 Dec 18. J Biol Chem. 2025. PMID: 39706275 Free PMC article.
-
An apical MRCK-driven morphogenetic pathway controls epithelial polarity.Nat Cell Biol. 2017 Sep;19(9):1049-1060. doi: 10.1038/ncb3592. Epub 2017 Aug 21. Nat Cell Biol. 2017. PMID: 28825699 Free PMC article.
-
Lgl, aPKC, and Crumbs regulate the Salvador/Warts/Hippo pathway through two distinct mechanisms.Curr Biol. 2010 Apr 13;20(7):573-81. doi: 10.1016/j.cub.2010.01.055. Epub 2010 Apr 1. Curr Biol. 2010. PMID: 20362447
-
PAR3-PAR6-atypical PKC polarity complex proteins in neuronal polarization.Cell Mol Life Sci. 2018 Aug;75(15):2735-2761. doi: 10.1007/s00018-018-2828-6. Epub 2018 Apr 25. Cell Mol Life Sci. 2018. PMID: 29696344 Free PMC article. Review.
-
Par-3 family proteins in cell polarity & adhesion.FEBS J. 2022 Feb;289(3):596-613. doi: 10.1111/febs.15754. Epub 2021 Mar 3. FEBS J. 2022. PMID: 33565714 Free PMC article. Review.
Cited by
-
Apical-basal polarity and the control of epithelial form and function.Nat Rev Mol Cell Biol. 2022 Aug;23(8):559-577. doi: 10.1038/s41580-022-00465-y. Epub 2022 Apr 19. Nat Rev Mol Cell Biol. 2022. PMID: 35440694 Review.
-
Signaling Pathways That Mediate Alveolar Macrophage Activation by Surfactant Protein A and IL-4.Front Immunol. 2022 Apr 4;13:860262. doi: 10.3389/fimmu.2022.860262. eCollection 2022. Front Immunol. 2022. PMID: 35444643 Free PMC article.
-
Distinct activities of Scrib module proteins organize epithelial polarity.Proc Natl Acad Sci U S A. 2020 May 26;117(21):11531-11540. doi: 10.1073/pnas.1918462117. Epub 2020 May 15. Proc Natl Acad Sci U S A. 2020. PMID: 32414916 Free PMC article.
-
The Drosophila neuroblast polarity cycle at a glance.J Cell Sci. 2024 Mar 1;137(5):jcs261789. doi: 10.1242/jcs.261789. Epub 2024 Mar 11. J Cell Sci. 2024. PMID: 38465513 Free PMC article.
-
Atypical Protein Kinase C Promotes its own Asymmetric Localisation by Phosphorylating Cdc42 in the C. elegans zygote.bioRxiv [Preprint]. 2024 Jun 14:2023.10.27.563985. doi: 10.1101/2023.10.27.563985. bioRxiv. 2024. PMID: 38009101 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
