Asexual Evolution and Forest Conditions Drive Genetic Parallelism in Phytophthora ramorum
- PMID: 32580470
- PMCID: PMC7357085
- DOI: 10.3390/microorganisms8060940
Asexual Evolution and Forest Conditions Drive Genetic Parallelism in Phytophthora ramorum
Abstract
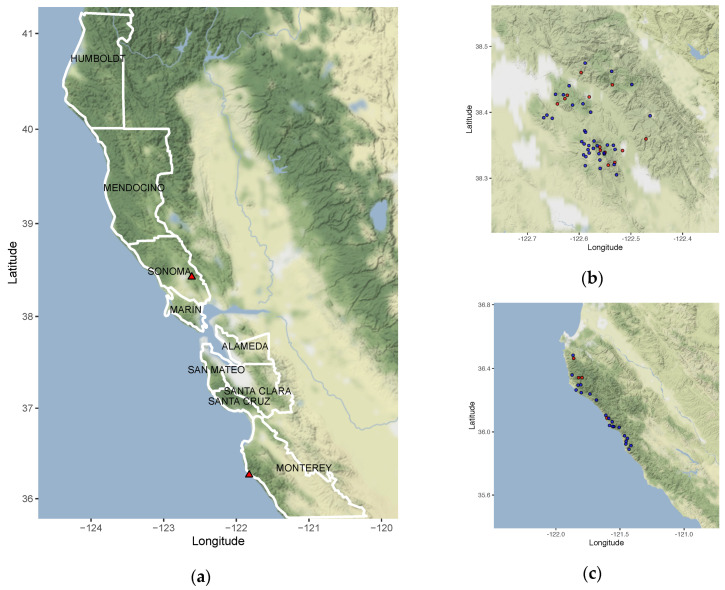
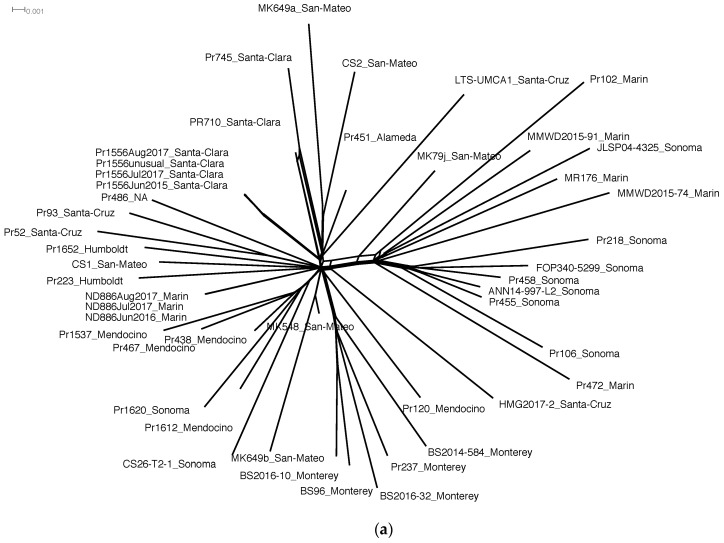
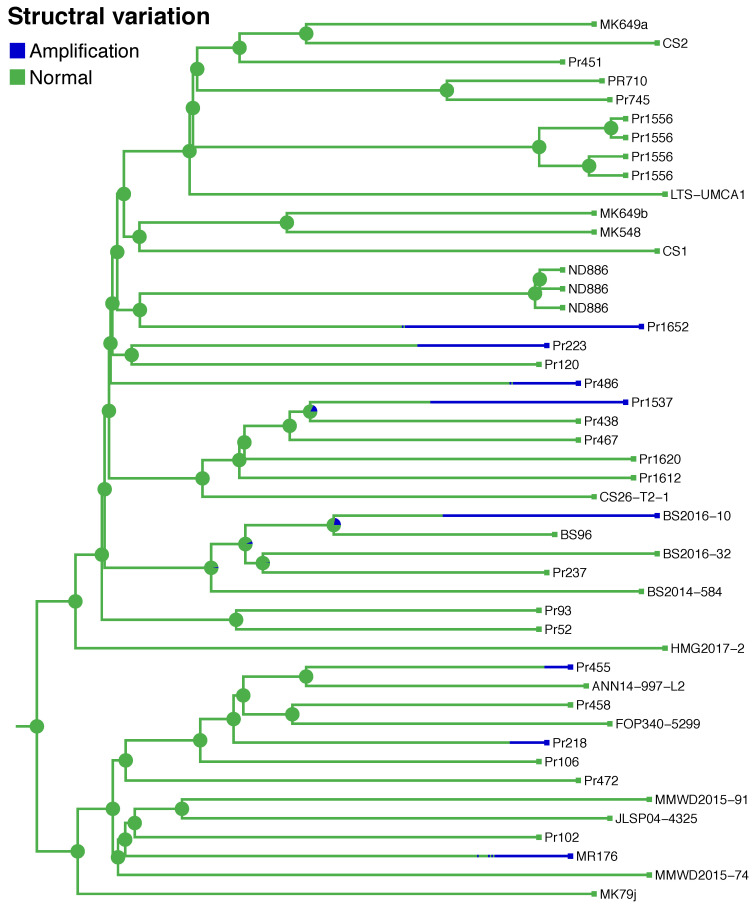
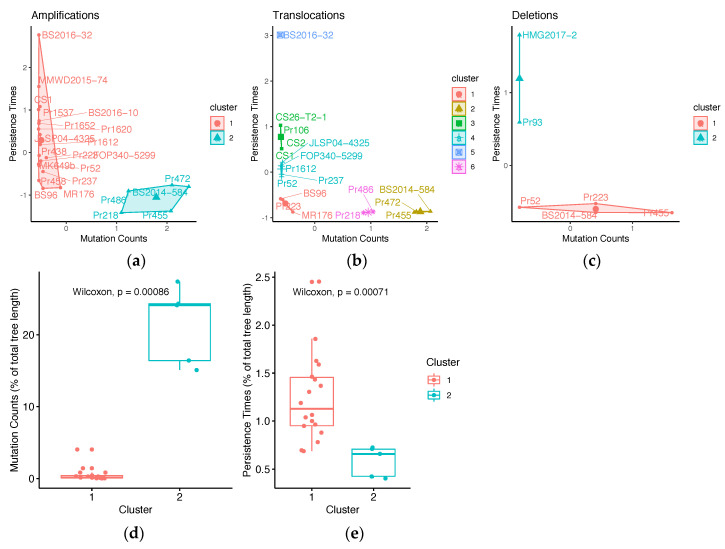
It is commonly assumed that asexual lineages are short-lived evolutionarily, yet many asexual organisms can generate genetic and phenotypic variation, providing an avenue for further evolution. Previous work on the asexual plant pathogen Phytophthora ramorum NA1 revealed considerable genetic variation in the form of Structural Variants (SVs). To better understand how SVs arise and their significance to the California NA1 population, we studied the evolutionary histories of SVs and the forest conditions associated with their emergence. Ancestral state reconstruction suggests that SVs arose by somatic mutations among multiple independent lineages, rather than by recombination. We asked if this unusual phenomenon of parallel evolution between isolated populations is transmitted to extant lineages and found that SVs persist longer in a population if their genetic background had a lower mutation load. Genetic parallelism was also found in geographically distant demes where forest conditions such as host density, solar radiation, and temperature, were similar. Parallel SVs overlap with genes involved in pathogenicity such as RXLRs and have the potential to change the course of an epidemic. By combining genomics and environmental data, we identified an unexpected pattern of repeated evolution in an asexual population and identified environmental factors potentially driving this phenomenon.
Keywords: Phytophthora ramorum; Structural Variants; asexual reproduction; forest pathology; parallel evolution.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures