

Movement-Related Signals in Sensory Areas: Roles in Natural Behavior
- PMID: 32580899
- PMCID: PMC8000520
- DOI: 10.1016/j.tins.2020.05.005
Movement-Related Signals in Sensory Areas: Roles in Natural Behavior
Abstract
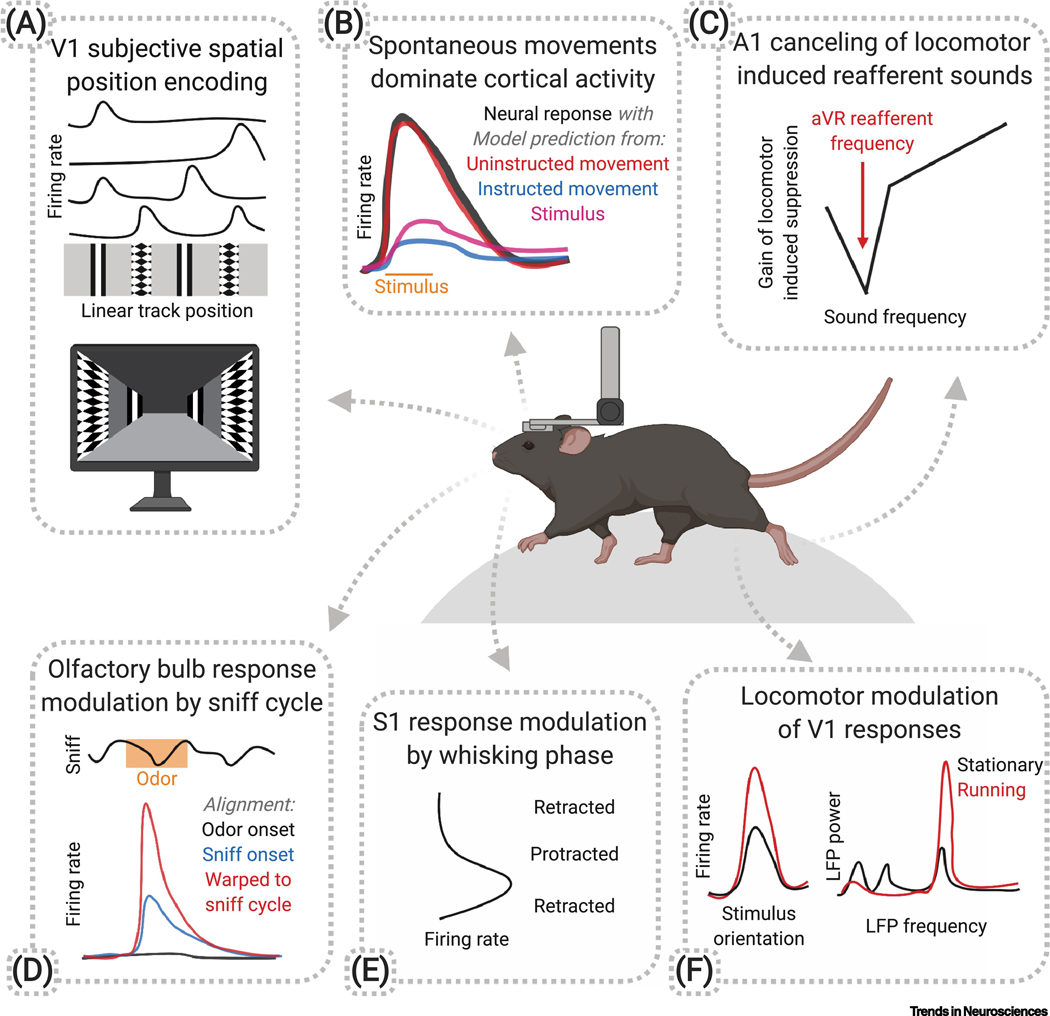
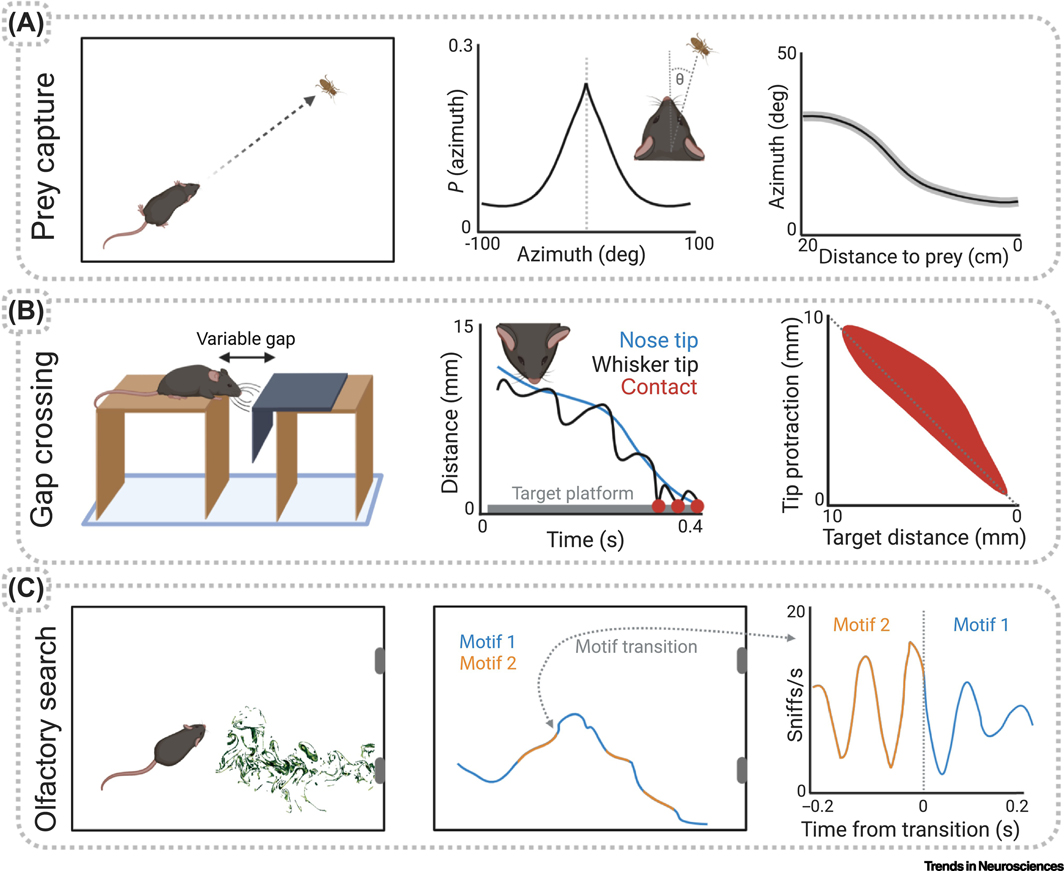
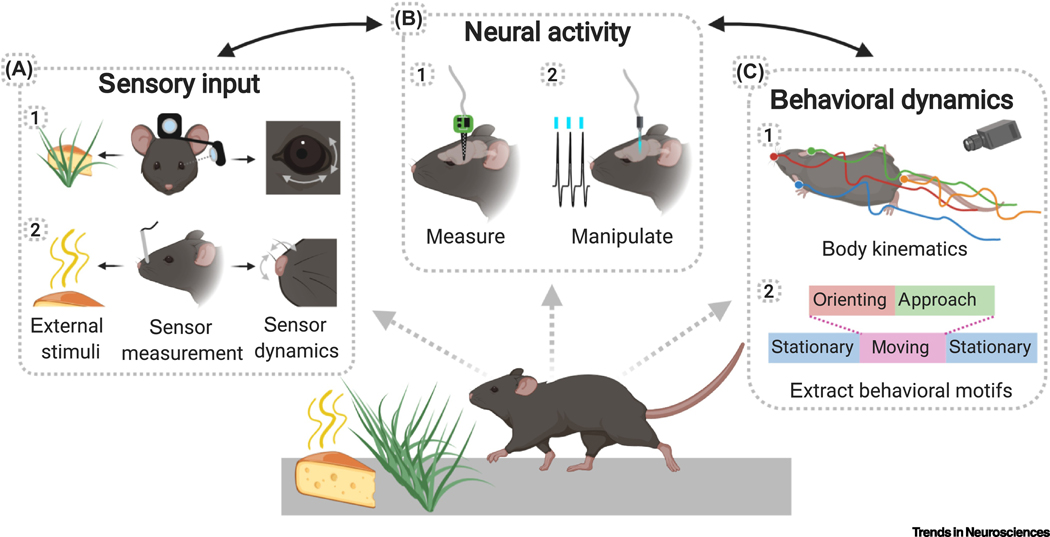
Recent studies have demonstrated prominent and widespread movement-related signals in the brain of head-fixed mice, even in primary sensory areas. However, it is still unknown what role these signals play in sensory processing. Why are these sensory areas 'contaminated' by movement signals? During natural behavior, animals actively acquire sensory information as they move through the environment and use this information to guide ongoing actions. In this context, movement-related signals could allow sensory systems to predict self-induced sensory changes and extract additional information about the environment. In this review we summarize recent findings on the presence of movement-related signals in sensory areas and discuss how their study, in the context of natural freely moving behaviors, could advance models of sensory processing.
Keywords: active sensation; cortical processing; ethology; sensory physiology.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
References
-
- Penfield W and Boldrey E. (1937) Somatic Motor And Sensory Representation In The Cerebral Cortex Of Man As Studied By Electrical Stimulation. Brain 60, 389–443
-
- Henschen SE (1893) On The Visual Path And Centre. Brain 16, 170–180
-
- Rust NC and Movshon JA (2005) In praise of artifice. Nat. Neurosci 8, 1647–1650 - PubMed
-
- Olshausen BA and Field DJ (2005) How close are we to understanding V1? Neural Comput. 17, 1665–1699 - PubMed
-
- Churchland Patricia S., Ramachandran Vilayanur S., and Sejnowski Terrence J. (1994) A critique of pure vision. In Large-scale neuronal theories of the brain (Koch Christof And Joel, ed), pp. 23, MIT Press
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
