

Reovirus Core Proteins λ1 and σ2 Promote Stability of Disassembly Intermediates and Influence Early Replication Events
- PMID: 32581098
- PMCID: PMC7431778
- DOI: 10.1128/JVI.00491-20
Reovirus Core Proteins λ1 and σ2 Promote Stability of Disassembly Intermediates and Influence Early Replication Events
Abstract
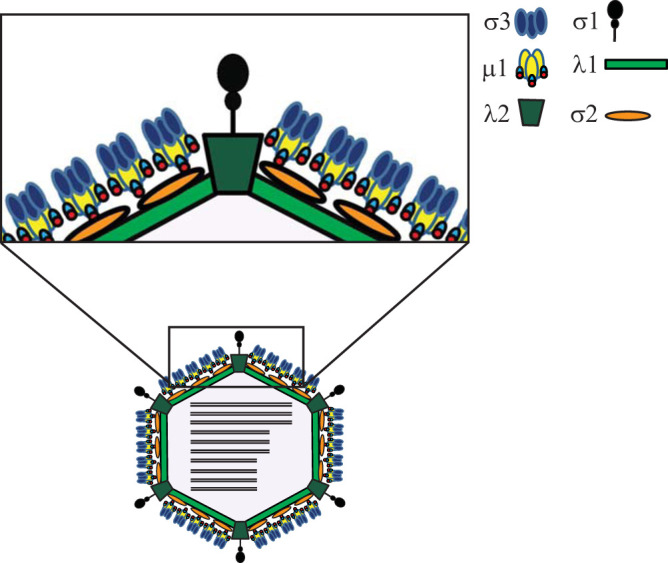
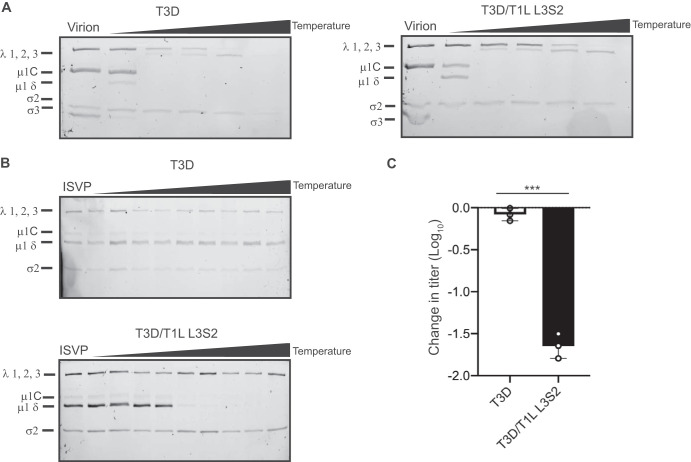
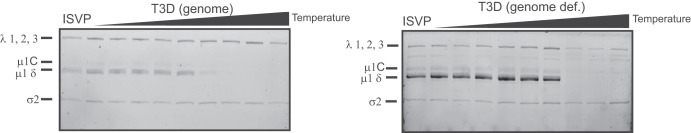
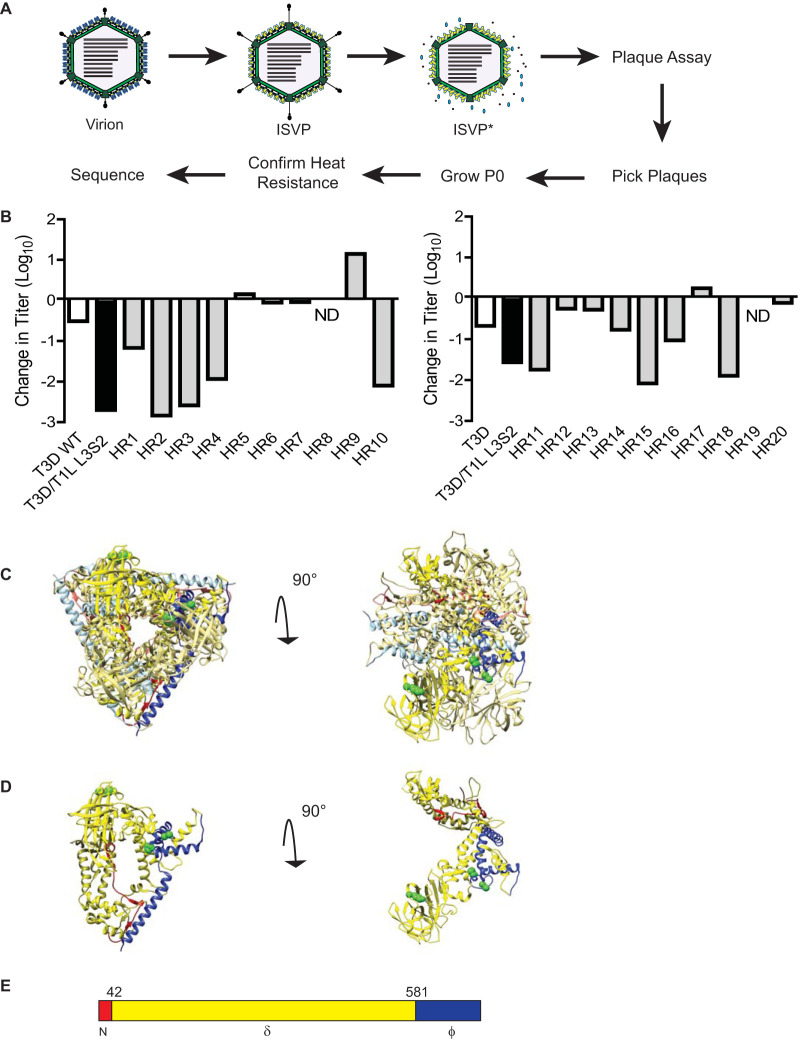
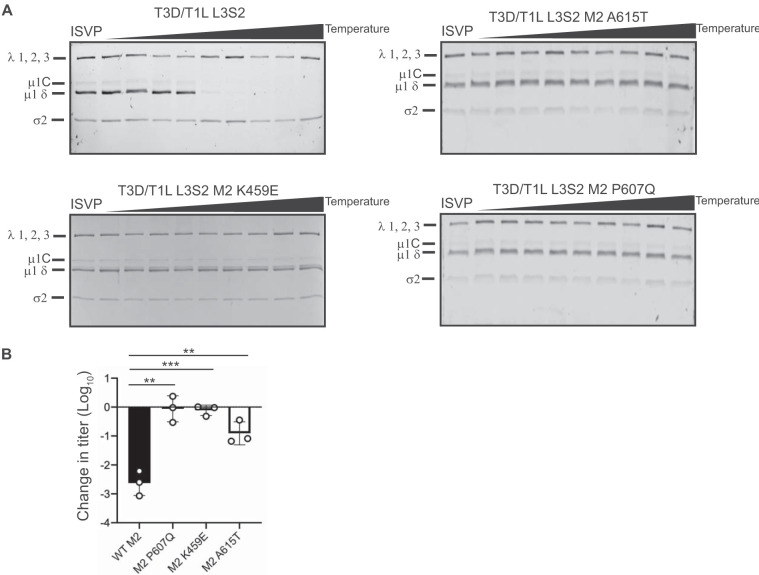
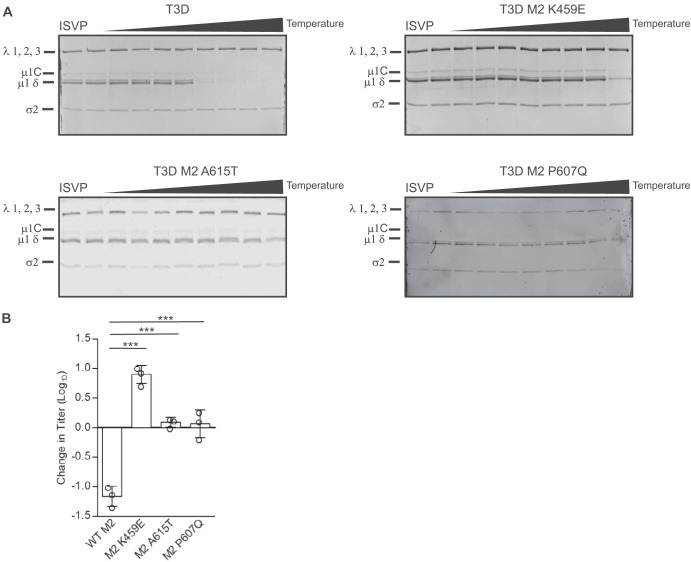
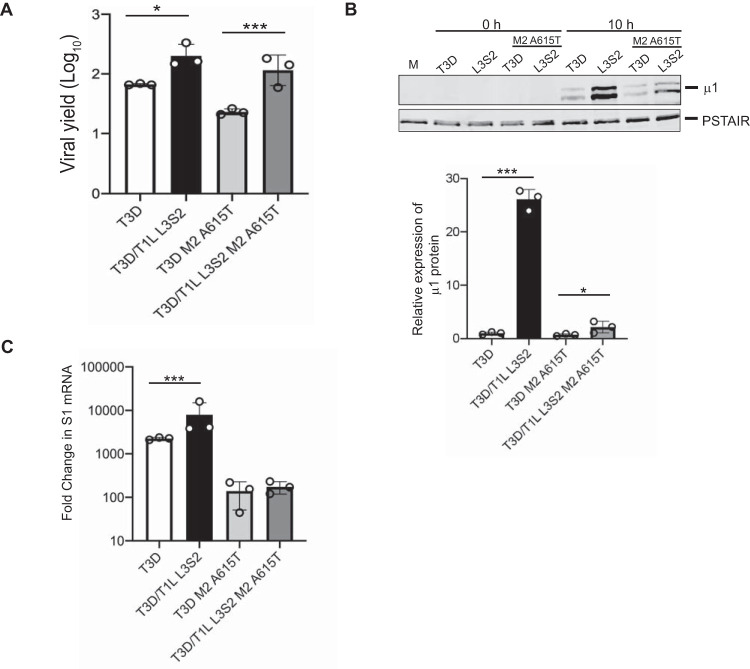
The capsids of mammalian reovirus contain two concentric protein shells, the core and the outer capsid. The outer capsid is composed of μ1-σ3 heterohexamers which surround the core. The core is composed of λ1 decamers held in place by σ2. After entry into the endosome, σ3 is proteolytically degraded and μ1 is cleaved and exposed to form infectious subvirion particles (ISVPs). ISVPs undergo further conformational changes to form ISVP*s, resulting in the release of μ1 peptides, which facilitate the penetration of the endosomal membrane to release transcriptionally active core particles into the cytoplasm. Previous work identified regions or specific residues within reovirus outer capsid proteins that impact the efficiency of cell entry. We examined the functions of the core proteins λ1 and σ2. We generated a reovirus T3D reassortant that carries strain T1L-derived σ2 and λ1 proteins (T3D/T1L L3S2). This virus displays lower ISVP stability and therefore converts to ISVP*s more readily. To identify the molecular basis for lability of T3D/T1L L3S2, we screened for hyperstable mutants of T3D/T1L L3S2 and identified three point mutations in μ1 that stabilize ISVPs. Two of these mutations are located in the C-terminal ϕ region of μ1, which has not previously been implicated in controlling ISVP stability. Independent of compromised ISVP stability, we also found that T3D/T1L L3S2 launches replication more efficiently and produces higher yields in infected cells than T3D. In addition to identifying a new role for the core proteins in disassembly events, these data highlight the possibility that core proteins may influence multiple stages of infection.IMPORTANCE Protein shells of viruses (capsids) have evolved to undergo specific changes to ensure the timely delivery of genetic material to host cells. The 2-layer capsid of reovirus provides a model system to study the interactions between capsid proteins and the changes they undergo during entry. We tested a virus in which the core proteins were derived from a different strain than the outer capsid. In comparison to the parental T3D strain, we found that this mismatched virus was less stable and completed conformational changes required for entry prematurely. Capsid stability was restored by introduction of specific changes to the outer capsid, indicating that an optimal fit between inner and outer shells maintains capsid function. Separate from this property, mismatch between these protein layers also impacted the capacity of the virus to initiate infection and produce progeny. This study reveals new insights into the roles of capsid proteins and their multiple functions during viral replication.
Keywords: capsid; double-stranded RNA virus; reovirus.
Copyright © 2020 American Society for Microbiology.
Figures
Similar articles
-
Components of the Reovirus Capsid Differentially Contribute to Stability.J Virol. 2019 Jan 4;93(2):e01894-18. doi: 10.1128/JVI.01894-18. Print 2019 Jan 15. J Virol. 2019. PMID: 30381491 Free PMC article.
-
Protein Mismatches Caused by Reassortment Influence Functions of the Reovirus Capsid.J Virol. 2018 Sep 26;92(20):e00858-18. doi: 10.1128/JVI.00858-18. Print 2018 Oct 15. J Virol. 2018. PMID: 30068646 Free PMC article.
-
The μ1 72-96 loop controls conformational transitions during reovirus cell entry.J Virol. 2013 Dec;87(24):13532-42. doi: 10.1128/JVI.01899-13. Epub 2013 Oct 2. J Virol. 2013. PMID: 24089575 Free PMC article.
-
Reovirus receptors, cell entry, and proapoptotic signaling.Adv Exp Med Biol. 2013;790:42-71. doi: 10.1007/978-1-4614-7651-1_3. Adv Exp Med Biol. 2013. PMID: 23884585 Free PMC article. Review.
-
From touchdown to transcription: the reovirus cell entry pathway.Curr Top Microbiol Immunol. 2010;343:91-119. doi: 10.1007/82_2010_32. Curr Top Microbiol Immunol. 2010. PMID: 20397070 Free PMC article. Review.
Cited by
-
The μ2 and λ1 Proteins of Mammalian Reovirus Modulate Early Events Leading to Induction of the Interferon Signaling Network.Viruses. 2022 Nov 26;14(12):2638. doi: 10.3390/v14122638. Viruses. 2022. PMID: 36560642 Free PMC article.
-
Molecular Characterization of Outer Capsid Proteins VP5 and VP7 of Grass Carp Reovirus.Viruses. 2022 May 12;14(5):1032. doi: 10.3390/v14051032. Viruses. 2022. PMID: 35632773 Free PMC article.
-
Mechanisms of Cell Entry by dsRNA Viruses: Insights for Efficient Delivery of dsRNA and Tools for Improved RNAi-Based Pest Control.Front Physiol. 2021 Nov 11;12:749387. doi: 10.3389/fphys.2021.749387. eCollection 2021. Front Physiol. 2021. PMID: 34858204 Free PMC article. Review.
-
Mammalian orthoreovirus capsid protein σ3 antagonizes RLR-mediated antiviral responses by degrading MAVS.mSphere. 2024 Jun 25;9(6):e0023624. doi: 10.1128/msphere.00236-24. Epub 2024 May 17. mSphere. 2024. PMID: 38757961 Free PMC article.
-
Approaches to Evaluating Necroptosis in Virus-Infected Cells.Subcell Biochem. 2023;106:37-75. doi: 10.1007/978-3-031-40086-5_2. Subcell Biochem. 2023. PMID: 38159223
References
-
- Dermody TS, Parker JSL, Sherry B. 2013. Orthoreoviruses, p 1304–1346. In Knipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed Lippincott Williams & Wilkins, Philadelphia, PA.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
