

Protein S-Nitrosylation of Human Cytomegalovirus pp71 Inhibits Its Ability To Limit STING Antiviral Responses
- PMID: 32581105
- PMCID: PMC7431796
- DOI: 10.1128/JVI.00033-20
Protein S-Nitrosylation of Human Cytomegalovirus pp71 Inhibits Its Ability To Limit STING Antiviral Responses
Abstract
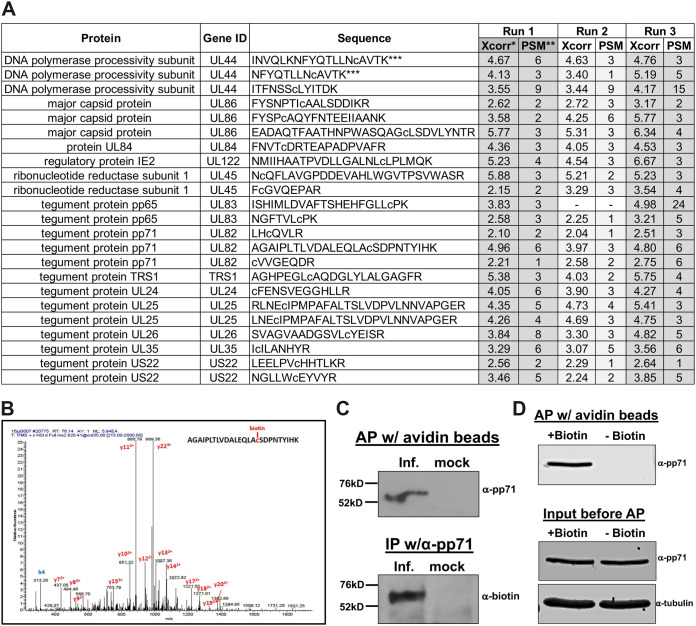
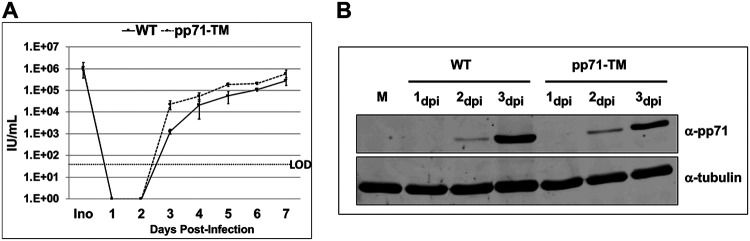
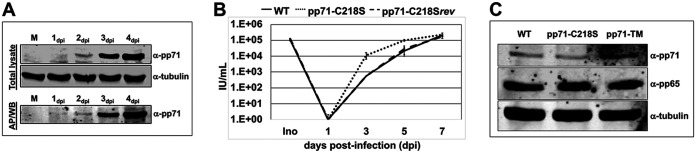
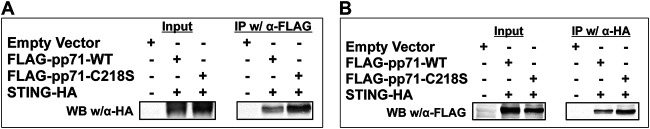
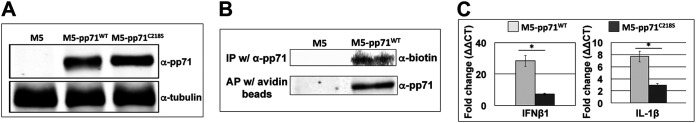
Human Cytomegalovirus (HCMV) is a ubiquitous pathogen that has coevolved with its host and, in doing so, is highly efficient in undermining antiviral responses that limit successful infections. As a result, HCMV infections are highly problematic in individuals with weakened or underdeveloped immune systems, including transplant recipients and newborns. Understanding how HCMV controls the microenvironment of an infected cell so as to favor productive replication is of critical importance. To this end, we took an unbiased proteomics approach to identify the highly reversible, stress-induced, posttranslational modification (PTM) protein S-nitrosylation on viral proteins to determine the biological impact on viral replication. We identified protein S-nitrosylation of 13 viral proteins during infection of highly permissive fibroblasts. One of these proteins, pp71, is critical for efficient viral replication, as it undermines host antiviral responses, including stimulator of interferon genes (STING) activation. By exploiting site-directed mutagenesis of the specific amino acids we identified in pp71 as protein S-nitrosylated, we found this pp71 PTM diminishes its ability to undermine antiviral responses induced by the STING pathway. Our results suggest a model in which protein S-nitrosylation may function as a host response to viral infection that limits viral spread.IMPORTANCE In order for a pathogen to establish a successful infection, it must undermine the host cell responses inhibitory to the pathogen. As such, herpesviruses encode multiple viral proteins that antagonize each host antiviral response, thereby allowing for efficient viral replication. Human Cytomegalovirus encodes several factors that limit host countermeasures to infection, including pp71. Herein, we identified a previously unreported posttranslational modification of pp71, protein S-nitrosylation. Using site-directed mutagenesis, we mutated the specific sites of this modification thereby blocking this pp71 posttranslational modification. In contexts where pp71 is not protein S-nitrosylated, host antiviral response was inhibited. The net result of this posttranslational modification is to render a viral protein with diminished abilities to block host responses to infection. This novel work supports a model in which protein S-nitrosylation may be an additional mechanism in which a cell inhibits a pathogen during the course of infection.
Keywords: HCMV; STING; UL82; cytomegalovirus; human herpesviruses; pp71; protein S-nitrosylation.
Copyright © 2020 American Society for Microbiology.
Figures
Similar articles
-
Protein-S-nitrosylation of human cytomegalovirus pp65 reduces its ability to undermine cGAS.J Virol. 2025 May 20;99(5):e0048125. doi: 10.1128/jvi.00481-25. Epub 2025 Apr 17. J Virol. 2025. PMID: 40243337 Free PMC article.
-
Human Cytomegalovirus Tegument Protein UL82 Inhibits STING-Mediated Signaling to Evade Antiviral Immunity.Cell Host Microbe. 2017 Feb 8;21(2):231-243. doi: 10.1016/j.chom.2017.01.001. Epub 2017 Jan 26. Cell Host Microbe. 2017. PMID: 28132838
-
Human Cytomegalovirus Utilizes Multiple Viral Proteins to Regulate the Basement Membrane Protein Nidogen 1.J Virol. 2022 Oct 26;96(20):e0133622. doi: 10.1128/jvi.01336-22. Epub 2022 Oct 11. J Virol. 2022. PMID: 36218358 Free PMC article.
-
Expanding the Known Functional Repertoire of the Human Cytomegalovirus pp71 Protein.Front Cell Infect Microbiol. 2020 Mar 12;10:95. doi: 10.3389/fcimb.2020.00095. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32226778 Free PMC article. Review.
-
Human cytomegalovirus tegument proteins (pp65, pp71, pp150, pp28).Virol J. 2012 Jan 17;9:22. doi: 10.1186/1743-422X-9-22. Virol J. 2012. PMID: 22251420 Free PMC article. Review.
Cited by
-
Universal STING Pathway-Activating Complexes Counteract Viral Immune Evasion and Boost Antiviral Responses.ACS Nano. 2025 Mar 4;19(8):8277-8293. doi: 10.1021/acsnano.4c18553. Epub 2025 Feb 23. ACS Nano. 2025. PMID: 39988898
-
Human Cytomegalovirus UL138 Protein Inhibits the STING Pathway and Reduces Interferon Beta mRNA Accumulation during Lytic and Latent Infections.mBio. 2021 Dec 21;12(6):e0226721. doi: 10.1128/mBio.02267-21. Epub 2021 Dec 14. mBio. 2021. PMID: 34903048 Free PMC article.
-
Innate Immune Responses to Herpesvirus Infection.Cells. 2021 Aug 18;10(8):2122. doi: 10.3390/cells10082122. Cells. 2021. PMID: 34440891 Free PMC article. Review.
-
Protein S-Nitrosylation: A Chemical Modification with Ubiquitous Biological Activities.Protein J. 2024 Aug;43(4):639-655. doi: 10.1007/s10930-024-10223-y. Epub 2024 Jul 28. Protein J. 2024. PMID: 39068633 Review.
-
Post-Translational Modifications of cGAS-STING: A Critical Switch for Immune Regulation.Cells. 2022 Sep 28;11(19):3043. doi: 10.3390/cells11193043. Cells. 2022. PMID: 36231006 Free PMC article. Review.
References
-
- McCormick AL, Mocarski ES Jr. 2007. Viral modulation of the host response to infection In Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, Whitley R, Yamanishi K (ed), Human herpesviruses: biology, therapy, and immunoprophylaxis. Cambridge University Press, Cambridge, United Kingdom. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
