

Accumulation of the RNA polymerase subunit RpoB depends on RNA editing by OsPPR16 and affects chloroplast development during early leaf development in rice
- PMID: 32583432
- PMCID: PMC7689822
- DOI: 10.1111/nph.16769
Accumulation of the RNA polymerase subunit RpoB depends on RNA editing by OsPPR16 and affects chloroplast development during early leaf development in rice
Abstract
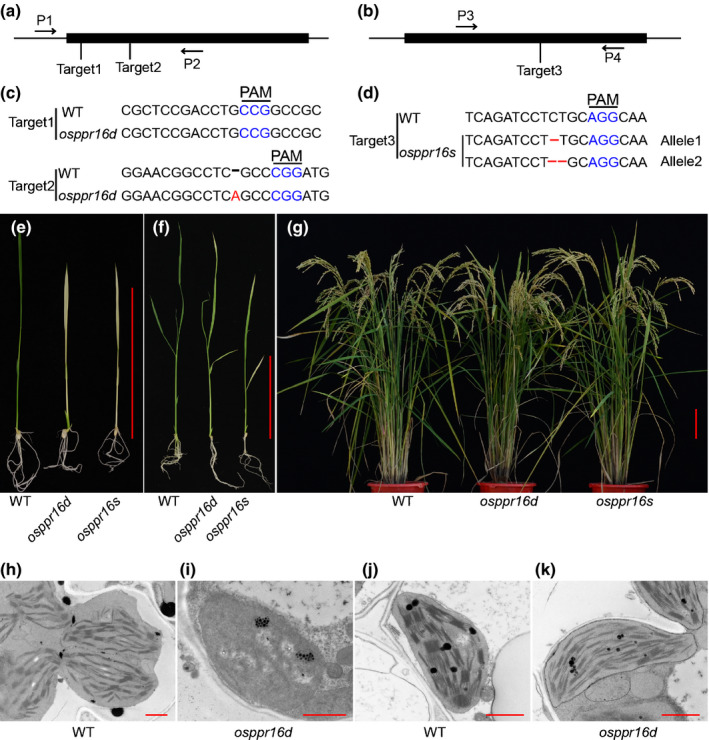
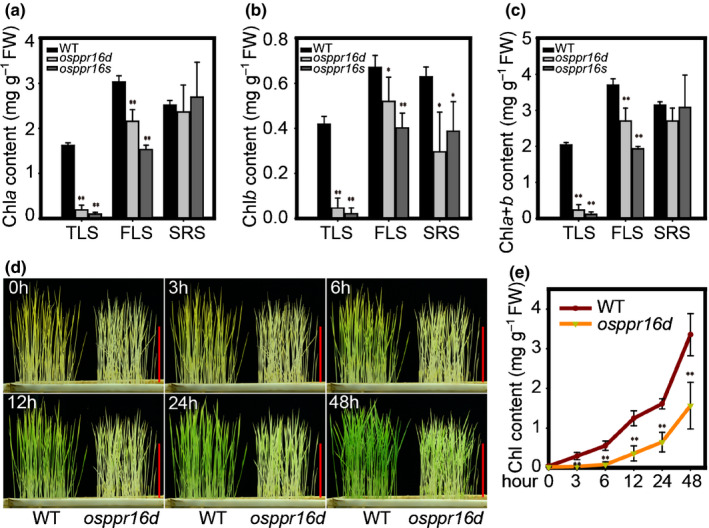
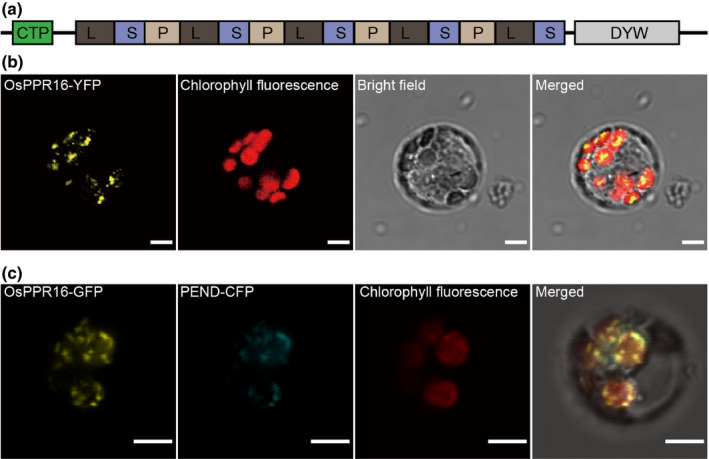
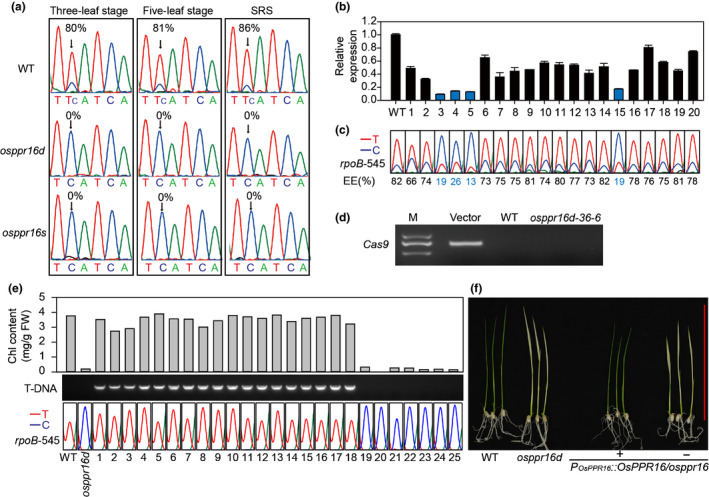
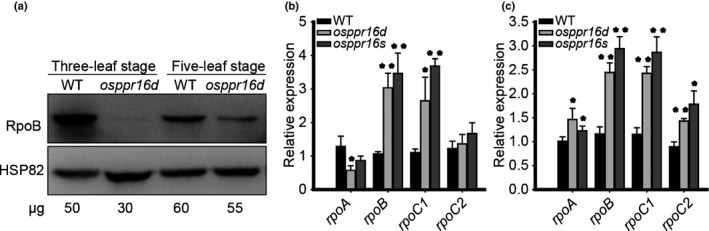
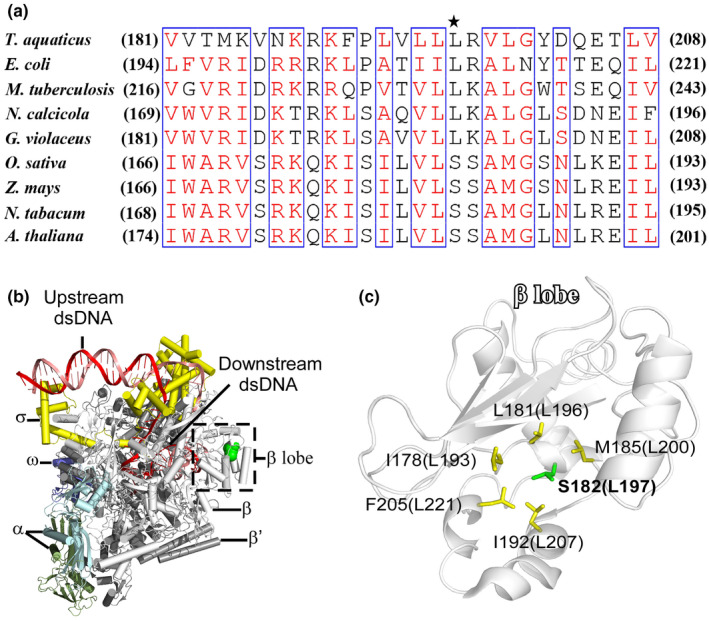
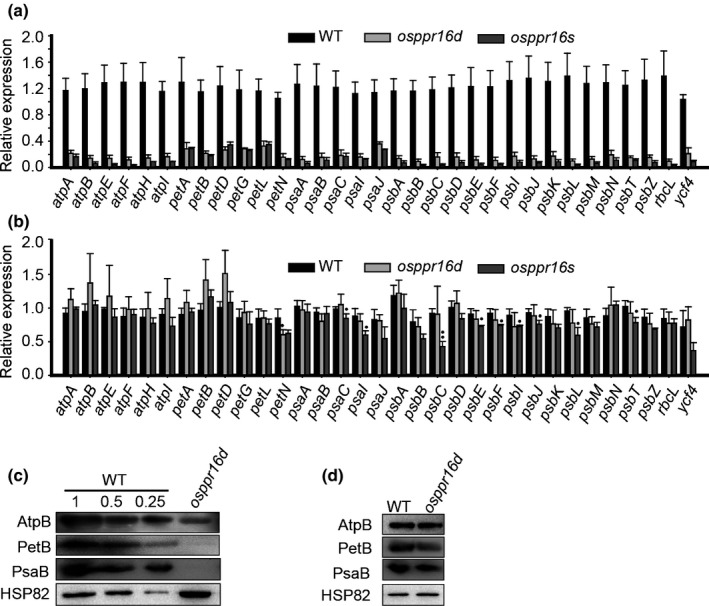
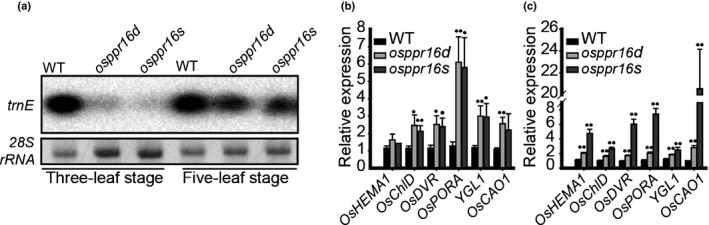
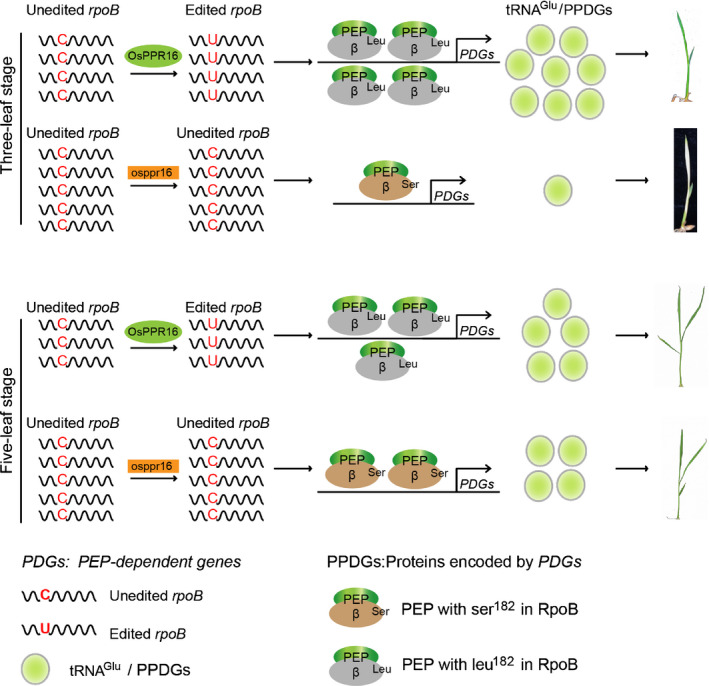
Plastid-encoded genes are coordinately transcribed by the nucleus-encoded RNA polymerase (NEP) and the plastid-encoded RNA polymerase (PEP). Resulting primary transcripts are frequently subject to RNA editing by cytidine-to-uridine conversions at specific sites. The physiological role of many editing events is largely unknown. Here, we have used the CRISPR/Cas9 technique in rice to knock out a member of the PLS-DYW subfamily of pentatricopeptide repeat (PPR) proteins. We found that OsPPR16 is responsible for a single editing event at position 545 in the chloroplast rpoB messenger RNA (mRNA), resulting in an amino acid change from serine to leucine in the β-subunit of the PEP. In striking contrast to loss-of-function mutations of the putative orthologue in Arabidopsis, which were reported to have no visible phenotype, knockout of OsPPR16 leads to impaired accumulation of RpoB, reduced expression of PEP-dependent genes, and a pale phenotype during early plant development. Thus, by editing the rpoB mRNA, OsPPR16 is required for faithful plastid transcription, which in turn is required for Chl synthesis and efficient chloroplast development. Our results provide new insights into the interconnection of the finely tuned regulatory mechanisms that operate at the transcriptional and post-transcriptional levels of plastid gene expression.
Keywords: Chl biosynthesis; RNA editing; RNA polymerase; chloroplast development; plastid; transcription.
©2020 The Authors. New Phytologist ©2020 New Phytologist Foundation.
Figures
References
-
- Asano T, Miyao A, Hirochika H, Kikuchi S, Kadowaki K‐i. 2013. A pentatricopeptide repeat gene of rice is required for splicing of chloroplast transcripts and RNA editing of ndhA . Plant Biotechnology 30: 57–64.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
