

Roles for a lipid phosphatase in the activation of its opposing lipid kinase
- PMID: 32583743
- PMCID: PMC7525815
- DOI: 10.1091/mbc.E18-09-0556
Roles for a lipid phosphatase in the activation of its opposing lipid kinase
Abstract
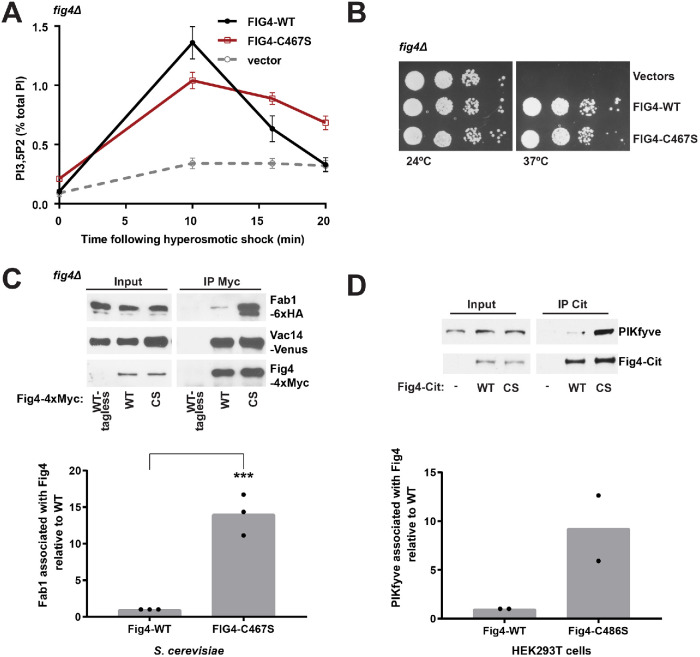
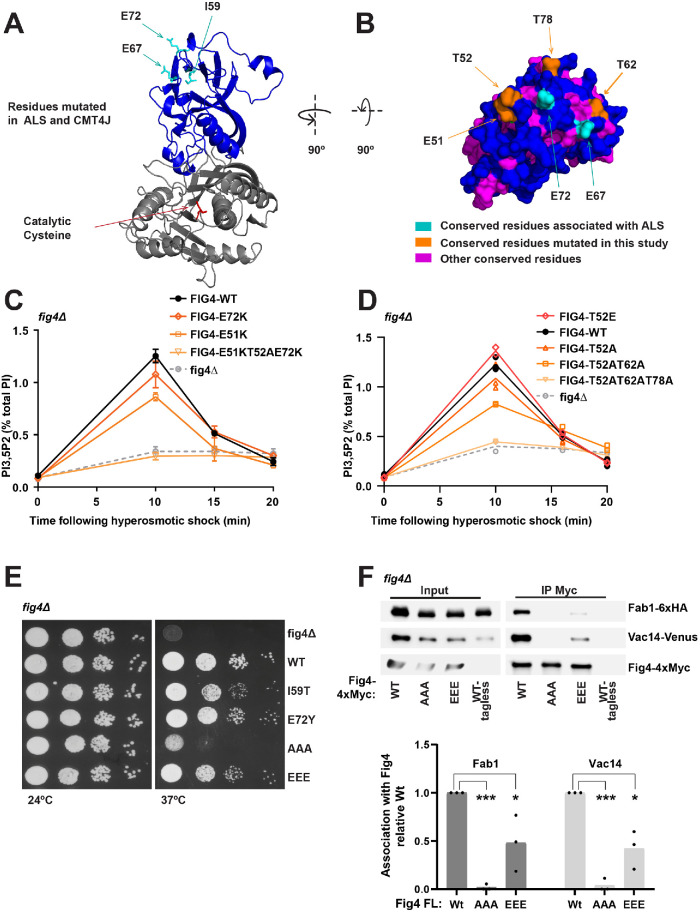
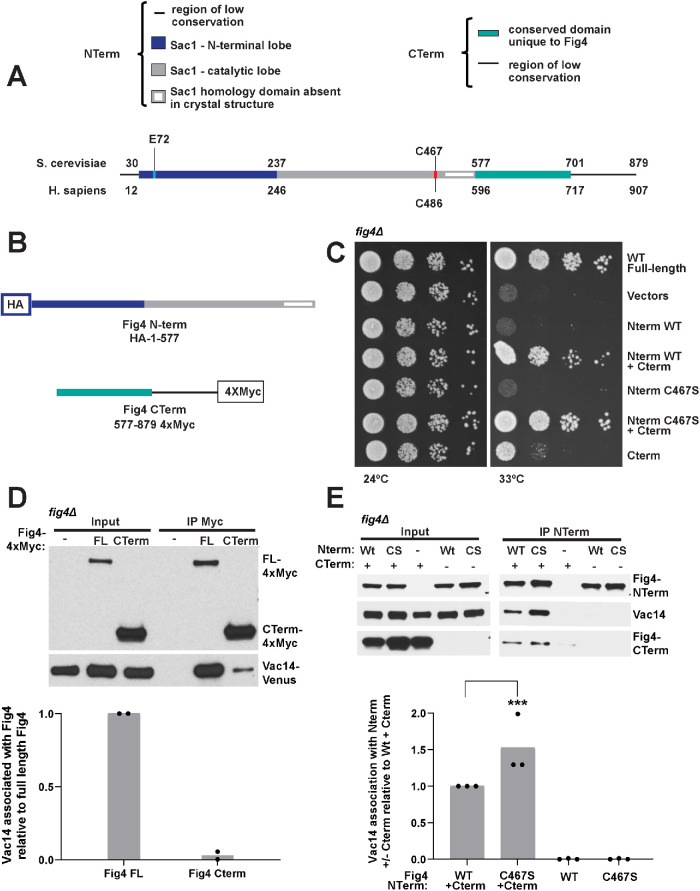
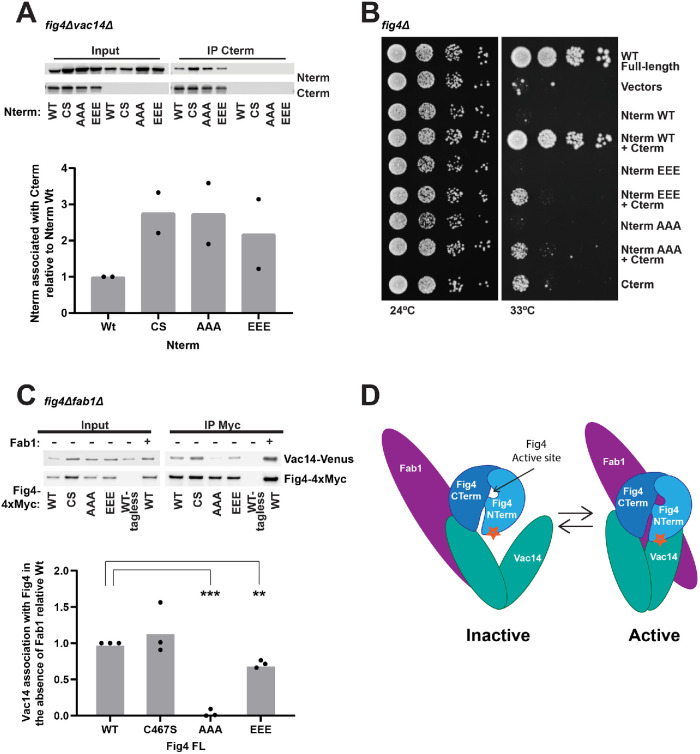
Fig4 is a phosphoinositide phosphatase that converts PI3,5P2 to PI3P. Paradoxically, mutation of Fig4 results in lower PI3,5P2, indicating that Fig4 is also required for PI3,5P2 production. Fig4 promotes elevation of PI3,5P2, in part, through stabilization of a protein complex that includes its opposing lipid kinase, Fab1, and the scaffold protein Vac14. Here we show that multiple regions of Fig4 contribute to its roles in the elevation of PI3,5P2: its catalytic site, an N-terminal disease-related surface, and a C-terminal region. We show that mutation of the Fig4 catalytic site enhances the formation of the Fab1-Vac14-Fig4 complex, and reduces the ability to elevate PI3,5P2. This suggests that independent of its lipid phosphatase function, the active site plays a role in the Fab1-Vac14-Fig4 complex. We also show that the N-terminal disease-related surface contributes to the elevation of PI3,5P2 and promotes Fig4 association with Vac14 in a manner that requires the Fig4 C-terminus. We find that the Fig4 C-terminus alone interacts with Vac14 in vivo and retains some functions of full-length Fig4. Thus, a subset of Fig4 functions are independent of its phosphatase domain and at least three regions of Fig4 play roles in the function of the Fab1-Vac14-Fig4 complex.
Figures
References
-
- Alonso A, Nunes-Xavier CE, Bayon Y, Pulido R. (2016). The extended family of protein tyrosine phosphatases. Methods Mol Biol , 1–23. - PubMed
-
- Berger P, Berger I, Schaffitzel C, Tersar K, Volkmer B, Suter U. (2006). Multi-level regulation of myotubularin-related protein-2 phosphatase activity by myotubularin-related protein-13/set-binding factor-2. Hum Mol Genet , 569–579. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
