

Exon-Skipping Oligonucleotides Restore Functional Collagen VI by Correcting a Common COL6A1 Mutation in Ullrich CMD
- PMID: 32585628
- PMCID: PMC7321786
- DOI: 10.1016/j.omtn.2020.05.029
Exon-Skipping Oligonucleotides Restore Functional Collagen VI by Correcting a Common COL6A1 Mutation in Ullrich CMD
Abstract
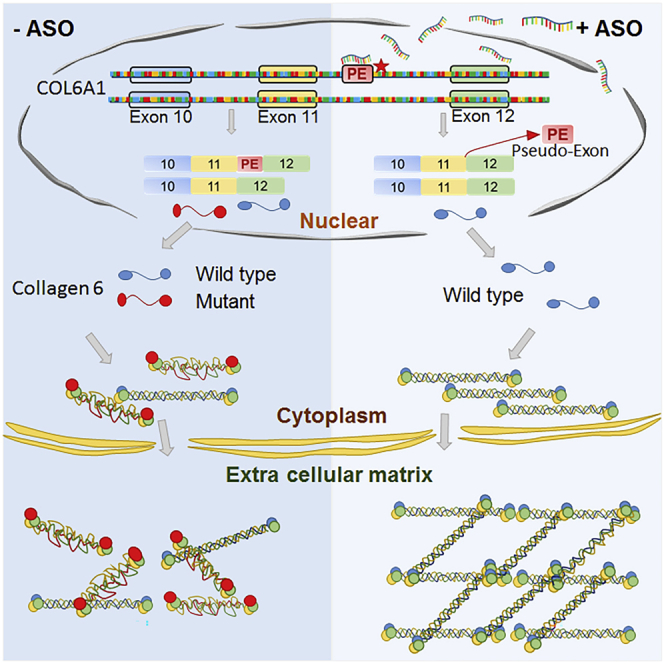
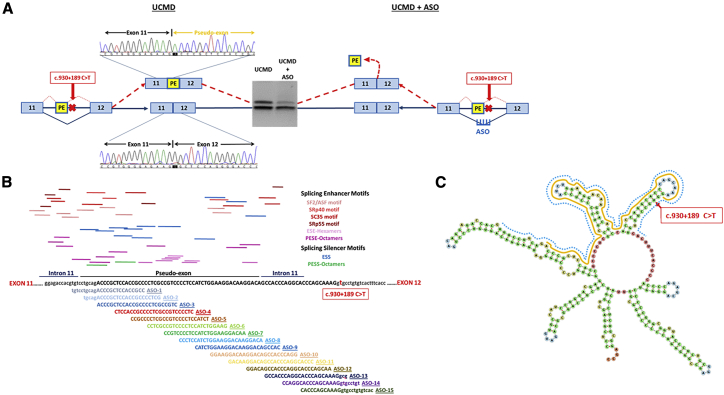
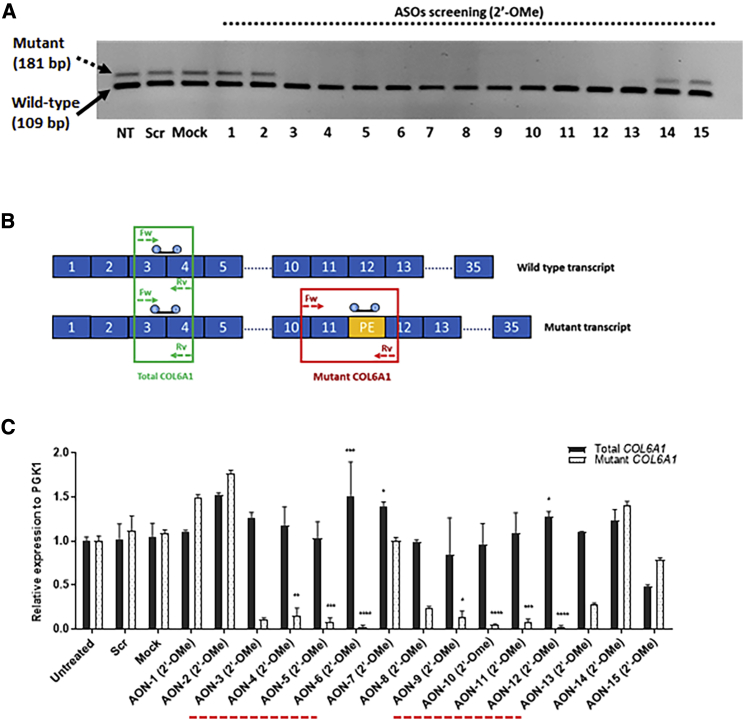
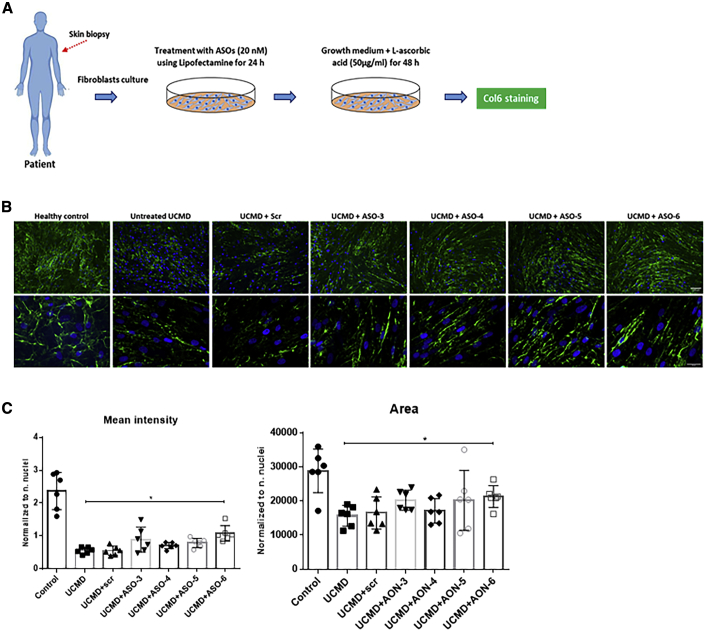
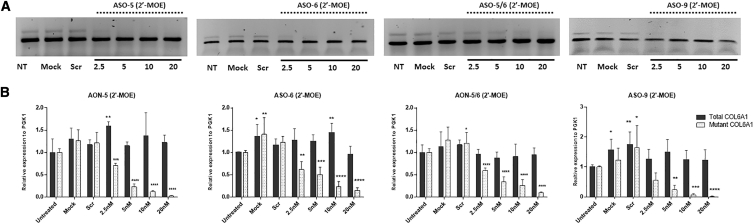
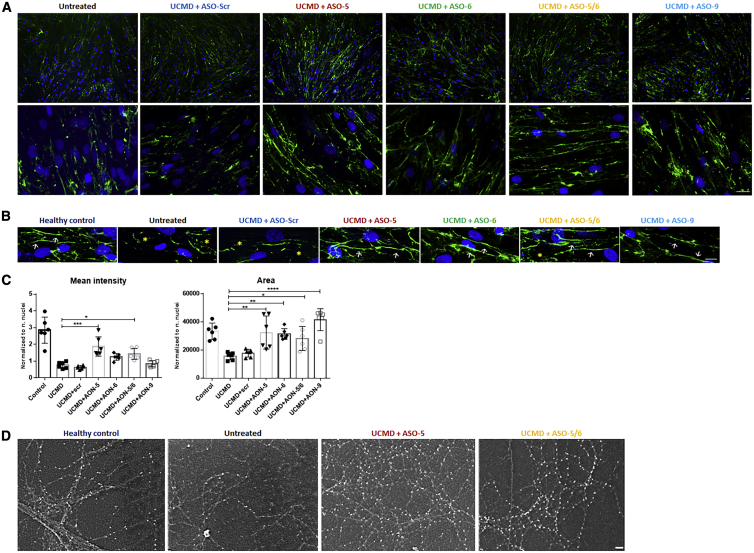
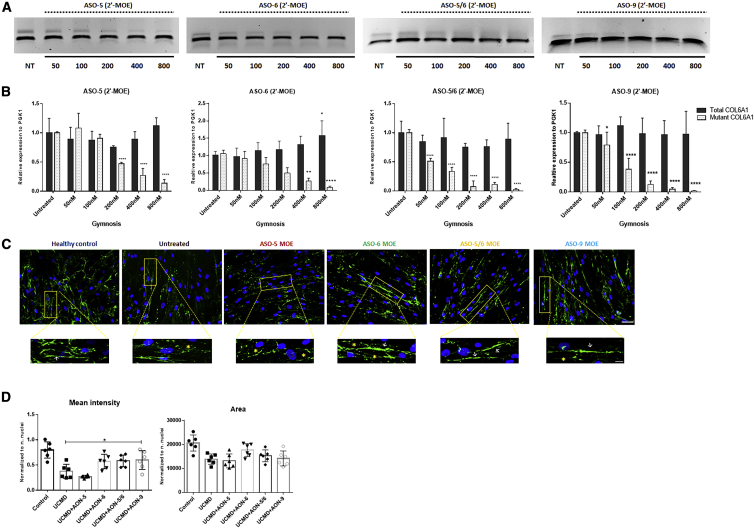
Collagen VI-related congenital muscular dystrophies (COL6-CMDs) are the second most common form of congenital muscular dystrophy. Currently, there is no effective treatment available. COL6-CMDs are caused by recessive or dominant mutations in one of the three genes encoding for the α chains of collagen type VI (COL6A1, COL6A2, and COL6A3). One of the most common mutations in COL6-CMD patients is a de novo deep intronic c.930+189C > T mutation in COL6A1 gene. This mutation creates a cryptic donor splice site and induces incorporation of a novel in-frame pseudo-exon in the mature transcripts. In this study, we systematically evaluated the splice switching approach using antisense oligonucleotides (ASOs) to correct this mutation. Fifteen ASOs were designed using the RNA-tiling approach to target the misspliced pseudo-exon and its flanking sequences. The efficiency of ASOs was evaluated at RNA, protein, and structural levels in skin fibroblasts established from four patients carrying the c.930+189C > T mutation. We identified two additional lead ASO candidates that efficiently induce pseudo-exon exclusion from the mature transcripts, thus allowing for the restoration of a functional collagen VI microfibrillar matrix. Our findings provide further evidence for ASO exon skipping as a therapeutic approach for COL6-CMD patients carrying this common intronic mutation.
Keywords: Collagen VI; Ullrich muscular dystrophy; antisense oligonucleotide; congenital muscular dystrophy; deep intronic mutation; exon skipping; extra cellular matrix.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Speer M.C., Tandan R., Rao P.N., Fries T., Stajich J.M., Bolhuis P.A., Jöbsis G.J., Vance J.M., Viles K.D., Sheffield K. Evidence for locus heterogeneity in the Bethlem myopathy and linkage to 2q37. Hum. Mol. Genet. 1996;5:1043–1046. - PubMed
-
- Clement E.M., Feng L., Mein R., Sewry C.A., Robb S.A., Manzur A.Y., Mercuri E., Godfrey C., Cullup T., Abbs S., Muntoni F. Relative frequency of congenital muscular dystrophy subtypes: analysis of the UK diagnostic service 2001-2008. Neuromuscul. Disord. 2012;22:522–527. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
