

Antifungal In Vitro Activity of Pilosulin- and Ponericin-Like Peptides from the Giant Ant Dinoponera quadriceps and Synergistic Effects with Antimycotic Drugs
- PMID: 32585881
- PMCID: PMC7344683
- DOI: 10.3390/antibiotics9060354
Antifungal In Vitro Activity of Pilosulin- and Ponericin-Like Peptides from the Giant Ant Dinoponera quadriceps and Synergistic Effects with Antimycotic Drugs
Abstract
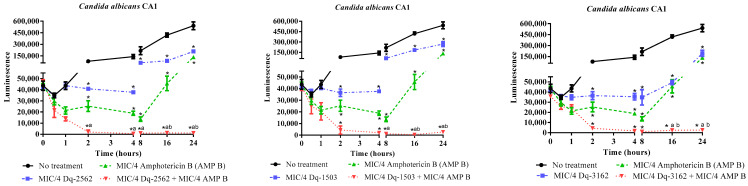
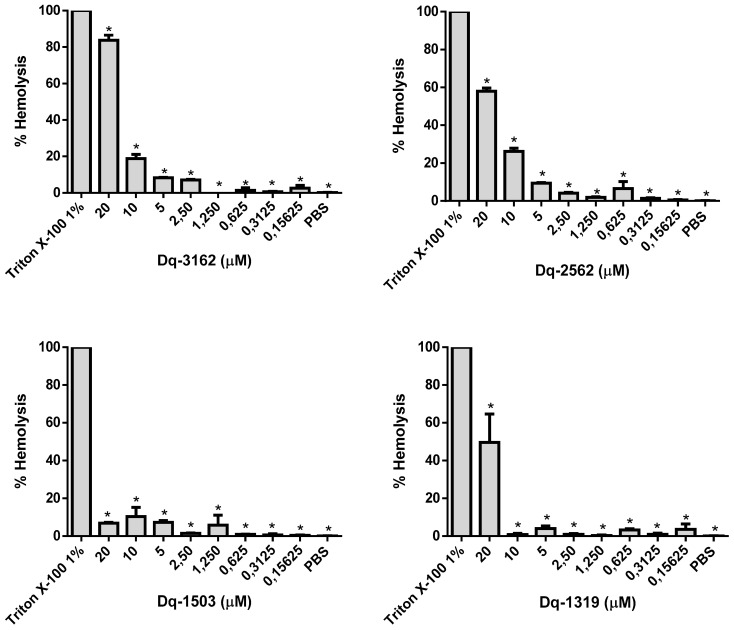
Venoms from ants comprise a rich source of bioactive peptides, including antimicrobial peptides. From the proteome and peptidome of the giant ant Dinoponera quadriceps venom, members of five known classes of antimicrobial peptides were disclosed (e.g., dermaseptin-, defensin-, ICK-, pilosulin- and ponericin-like types). Based on comparative analysis, these family members have structural determinants that indicate they could display antimicrobial activities. In previous works, pilosulin- and ponericin-like peptides were demonstrated to be active against bacteria, fungi, and parasites. Herein, the antifungal activity of ponericin- and pilosulin-like peptides were assessed, aiming at the expansion of the knowledge about AMPs in predatory ants and the development of new microbicide strategies to deal with difficult-to-treat fungal infections. Synthetic pilosulin- (Dq-2562, Dq-1503, and Dq-1319) and ponericin-like (Dq-3162) peptides were evaluated for their fungicide and fungistatic activities against different species of Candida, including a drug-resistant clinical strain. The MICs and MLCs were determined for all peptides individually and in combination with general antifungal drugs by the microdilution method. The time-kill kinetic curves were set up by means of a luminescent reagent, of which the light signal is proportional to the number of viable cells. The candicidal synergism observed by the combination of subinhibitory concentrations of peptides and general antimycotic drugs were quantified by the checkerboard test and fluorescent dye permeation assay. The influence of ergosterol on the antifungal activity was verified by supplementation of culture medium. The pilosulin- (Dq-2562 and Dq-1503) and ponericin-like (Dq-3162) were the most active peptides, displaying a broad spectrum of antifungal activity in vitro, with MICs in the range of 0.625 to 10 µM. The combination of peptides and conventional antimycotic drugs displayed a synergistic reduction in the MIC values of individual peptides and drugs, while soluble ergosterol in the culture medium increased the MICs. The fungicide and fungistatic activity of the individual peptides and peptides in combination with antimycotics were time-dependent with a rapid onset of action and long-lasting effect, which involved membrane disruption as an underlying mechanism of their action. Altogether, pilosulin- and ponericin-like peptides from the giant ant D. quadriceps venom display a broad-spectrum of candicidal activity, what allows their inclusion in the row of the antifungal peptides and gives support for further studies on the development of strategies to fight candidiasis.
Keywords: Dinoponera quadriceps; antifungal peptides; antimicrobial peptides; combination drugs; drug-resistant yeasts; fungicidal synergism; giant ant venom-peptides; pilosulin-like peptides; ponericin-like peptides.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study.
Figures




Similar articles
-
Antimicrobial activity of synthetic Dq-3162, a 28-residue ponericin G-like dinoponeratoxin from the giant ant Dinoponera quadriceps venom, against carbapenem-resistant bacteria.Toxicon. 2020 Nov;187:19-28. doi: 10.1016/j.toxicon.2020.08.015. Epub 2020 Aug 27. Toxicon. 2020. PMID: 32861765
-
Comprehensive analysis of peptides and low molecular weight components of the giant ant Dinoponera quadriceps venom.Biol Chem. 2020 Jul 28;401(8):945-954. doi: 10.1515/hsz-2019-0397. Biol Chem. 2020. PMID: 32229648
-
Comprehensive analysis of peptides and low molecular weight components of the giant ant Dinoponera quadriceps venom.Biol Chem. 2019 Dec 1:/j/bchm.just-accepted/hsz-2019-397ja-01/hsz-2019-397ja-01.xml. doi: 10.1515/hsz-2019-397ja-01. Online ahead of print. Biol Chem. 2019. PMID: 32087061
-
Pilosulins: a review of the structure and mode of action of venom peptides from an Australian ant Myrmecia pilosula.Toxicon. 2015 May;98:54-61. doi: 10.1016/j.toxicon.2015.02.013. Epub 2015 Feb 25. Toxicon. 2015. PMID: 25725257 Review.
-
In vitro antifungal activities of anidulafungin and micafungin, licensed agents and the investigational triazole posaconazole as determined by NCCLS methods for 12,052 fungal isolates: review of the literature.Rev Iberoam Micol. 2003 Dec;20(4):121-36. Rev Iberoam Micol. 2003. PMID: 15456349 Review.
Cited by
-
Exploring the frontiers of therapeutic breadth of antifungal peptides: A new avenue in antifungal drugs.J Ind Microbiol Biotechnol. 2024 Jan 9;51:kuae018. doi: 10.1093/jimb/kuae018. J Ind Microbiol Biotechnol. 2024. PMID: 38710584 Free PMC article. Review.
-
Multipurpose peptides: The venoms of Amazonian stinging ants contain anthelmintic ponericins with diverse predatory and defensive activities.Biochem Pharmacol. 2021 Oct;192:114693. doi: 10.1016/j.bcp.2021.114693. Epub 2021 Jul 22. Biochem Pharmacol. 2021. PMID: 34302796 Free PMC article.
-
Antifungal activity of protein hydrolysates from Thai Phatthalung Sangyod rice (Oryza sativa L.) seeds.Vet World. 2023 May;16(5):1018-1028. doi: 10.14202/vetworld.2023.1018-1028. Epub 2023 May 13. Vet World. 2023. PMID: 37576760 Free PMC article.
-
Unravelling the potential of insects for medicinal purposes - A comprehensive review.Heliyon. 2023 Apr 29;9(5):e15938. doi: 10.1016/j.heliyon.2023.e15938. eCollection 2023 May. Heliyon. 2023. PMID: 37206028 Free PMC article. Review.
-
Antifungal Polymeric Materials and Nanocomposites.Front Bioeng Biotechnol. 2021 Dec 24;9:780328. doi: 10.3389/fbioe.2021.780328. eCollection 2021. Front Bioeng Biotechnol. 2021. PMID: 35004642 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
