

Lysophosphatidylcholine acyltransferase 2 (LPCAT2) co-localises with TLR4 and regulates macrophage inflammatory gene expression in response to LPS
- PMID: 32587324
- PMCID: PMC7316826
- DOI: 10.1038/s41598-020-67000-x
Lysophosphatidylcholine acyltransferase 2 (LPCAT2) co-localises with TLR4 and regulates macrophage inflammatory gene expression in response to LPS
Abstract
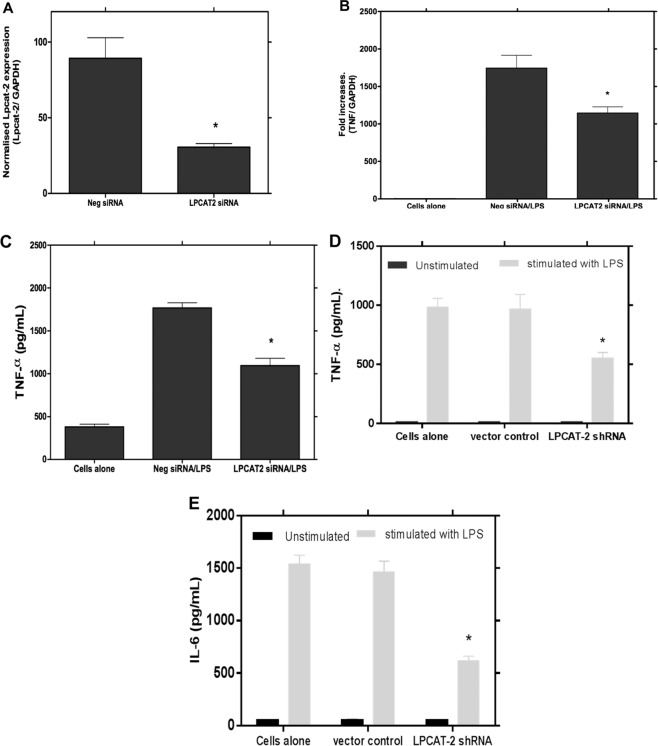
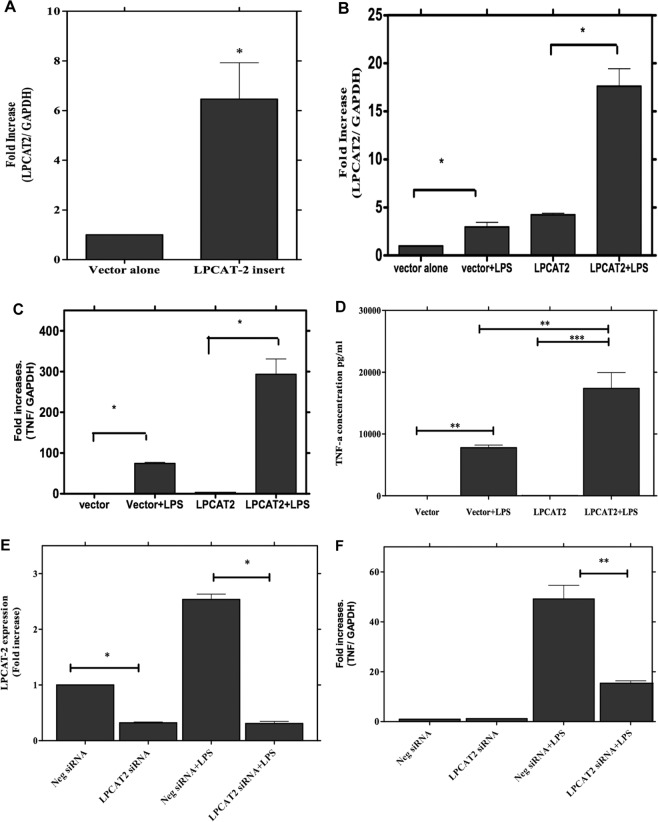
Despite extensive investigations, an effective treatment for sepsis remains elusive and a better understanding of the inflammatory response to infection is required to identify potential new targets for therapy. In this study we have used RNAi technology to show, for the first time, that the inducible lysophosphatidylcholine acyltransferase 2 (LPCAT2) plays a key role in macrophage inflammatory gene expression in response to stimulation with bacterial ligands. Using siRNA- or shRNA-mediated knockdown, we demonstrate that, in contrast to the constitutive LPCAT1, LPCAT2 is required for macrophage cytokine gene expression and release in response to TLR4 and TLR2 ligand stimulation but not for TLR-independent stimuli. In addition, cells transfected to overexpress LPCAT2 exhibited increased expression of inflammatory genes in response to LPS and other bacterial ligands. Furthermore, we have used immunoprecipitation and Western blotting to show that in response to LPS, LPCAT2, but not LPCAT1, rapidly associates with TLR4 and translocates to membrane lipid raft domains. Our data thus suggest a novel mechanism for the regulation of inflammatory gene expression in response to bacterial stimuli and highlight LPCAT2 as a potential therapeutic target for development of anti-inflammatory and anti-sepsis therapies.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- van der Poll T, van Deventer SJ. Cytokines and anticytokines in the pathogenesis of sepsis. Infectious disease clinics of North America. 1999;13(413–426):ix.. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
