

Enhancing fatty acid oxidation negatively regulates PPARs signaling in the heart
- PMID: 32592696
- PMCID: PMC7494556
- DOI: 10.1016/j.yjmcc.2020.06.008
Enhancing fatty acid oxidation negatively regulates PPARs signaling in the heart
Abstract
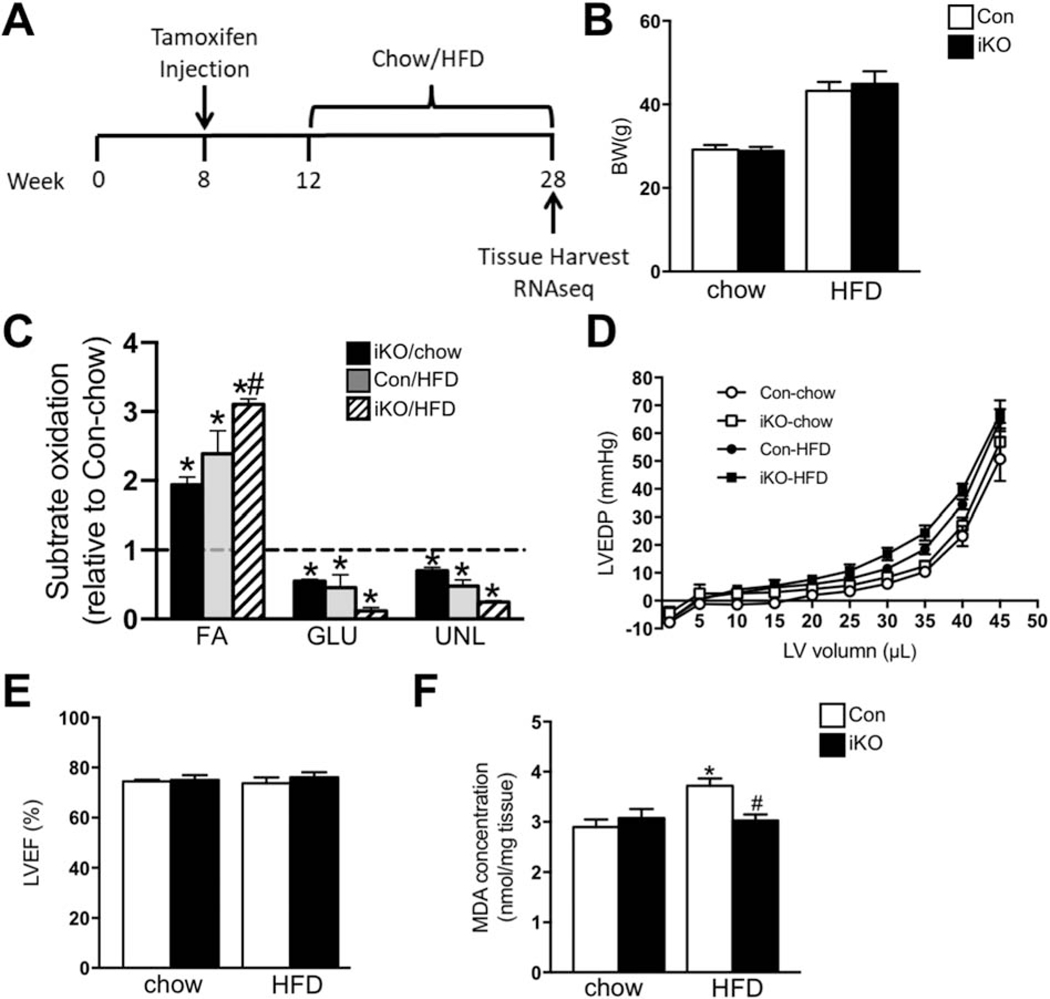
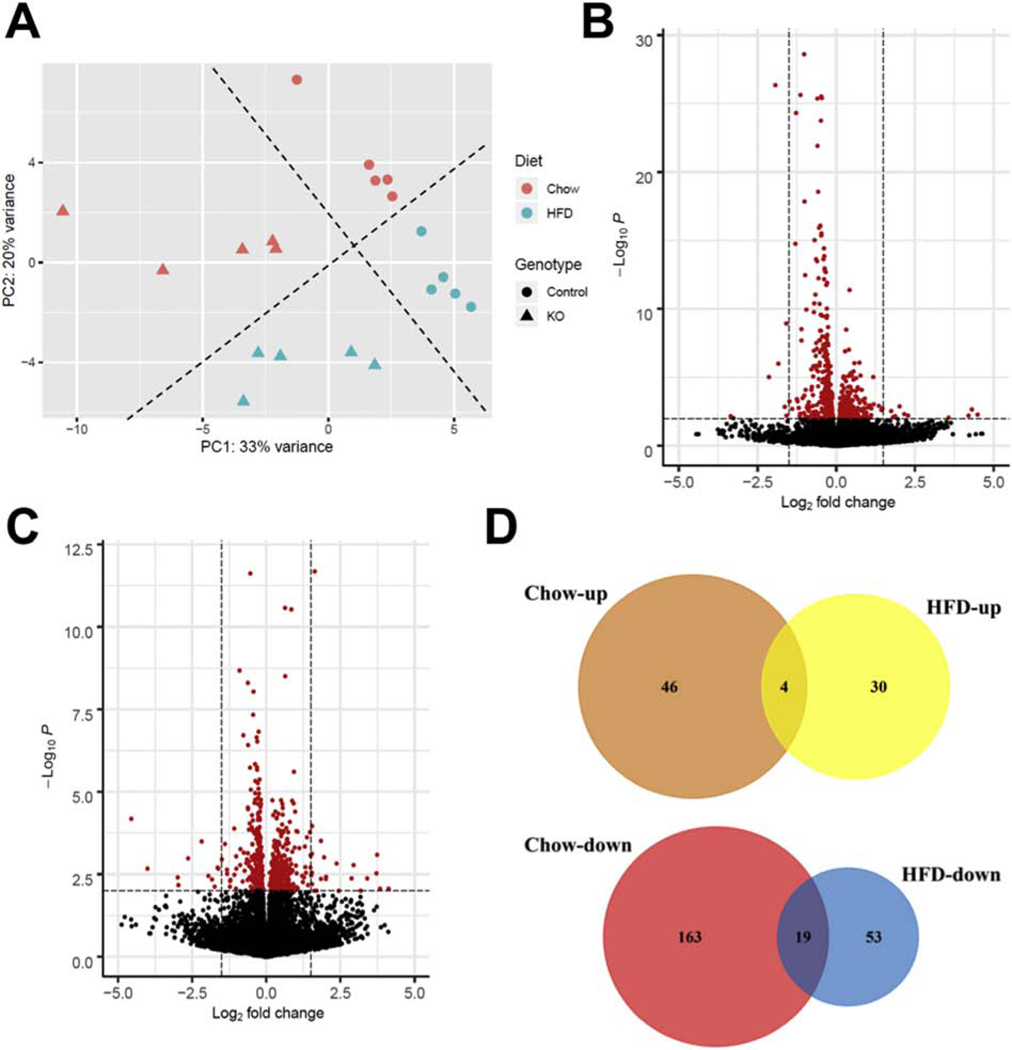
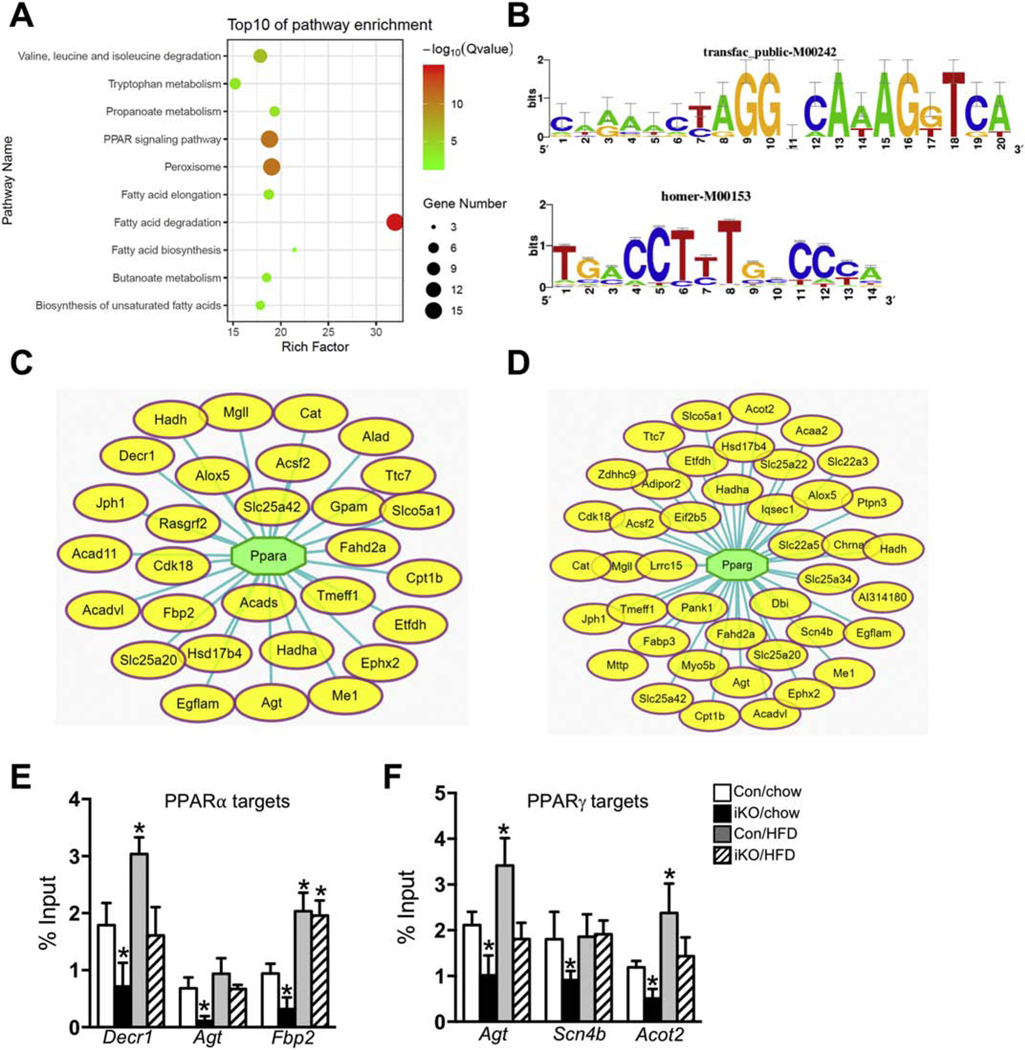
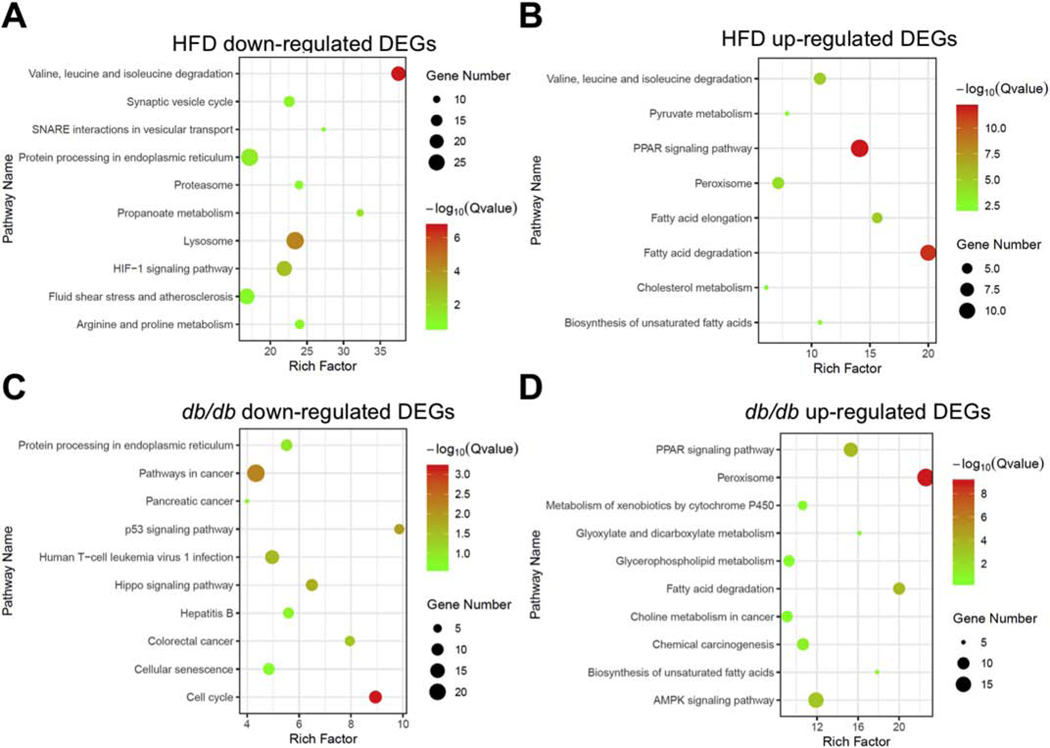
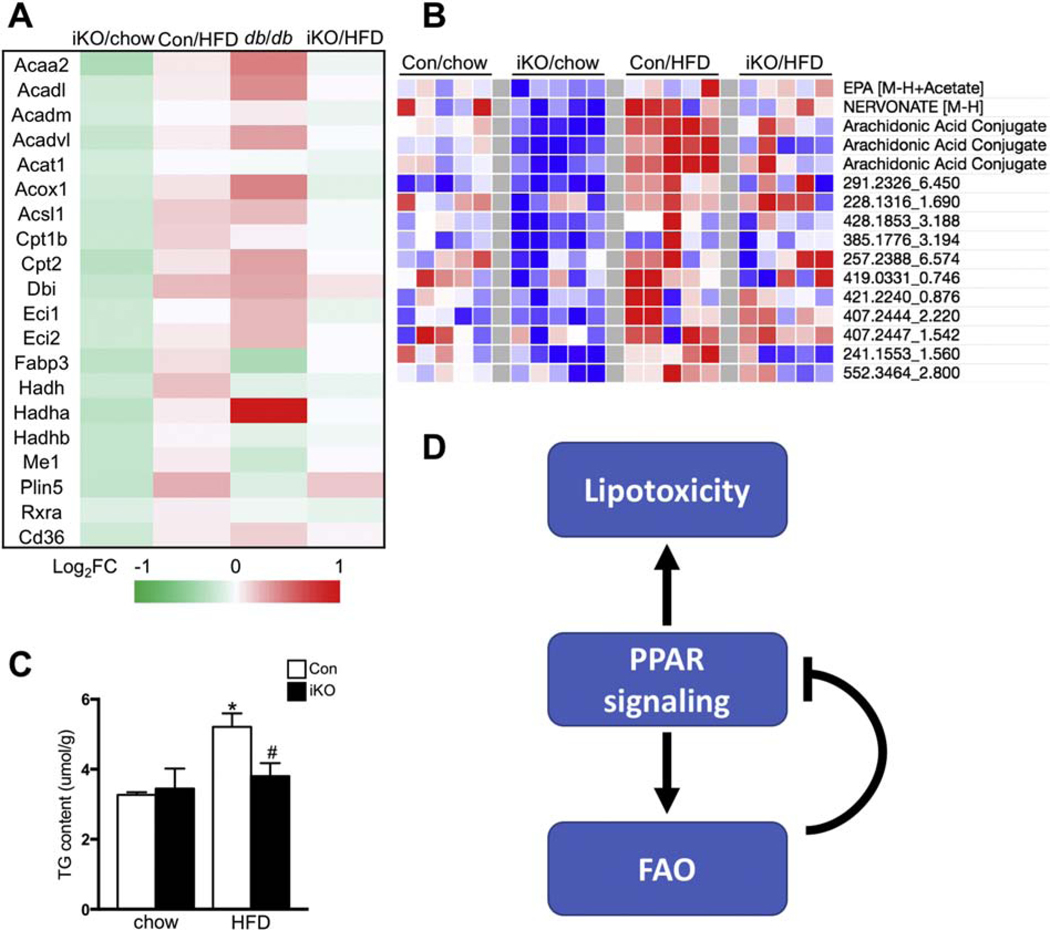
High fatty acid oxidation (FAO) is associated with lipotoxicity, but whether it causes lipotoxic cardiomyopathy remains controversial. Molecular mechanisms that may be responsible for FAO-induced lipotoxic cardiomyopathy are also elusive. In this study, increasing FAO by genetic deletion of acetyl-CoA carboxylase 2 (ACC2) did not induce cardiac dysfunction after 16 weeks of high fat diet (HFD) feeding. This suggests that increasing FAO, per se, does not cause metabolic cardiomyopathy in obese mice. We compared transcriptomes of control and ACC2 deficient mouse hearts under chow- or HFD-fed conditions. ACC2 deletion had a significant impact on the global transcriptome including downregulation of the peroxisome proliferator-activated receptors (PPARs) signaling and fatty acid degradation pathways. Increasing fatty acids by HFD feeding normalized expression of fatty acid degradation genes in ACC2 deficient mouse hearts to the same level as the control mice. In contrast, cardiac transcriptome analysis of the lipotoxic mouse model (db/db) showed an upregulation of PPARs signaling and fatty acid degradation pathways. Our results suggest that enhancing FAO by genetic deletion of ACC2 negatively regulates PPARs signaling through depleting endogenous PPAR ligands, which can serve as a negative feedback mechanism to prevent excess activation of PPAR signaling under non-obese condition. In obesity, excessive lipid availability negates the feedback mechanism resulting in over activation of PPAR cascade, thus contributes to the development of cardiac lipotoxicity.
Keywords: Fatty acid oxidation; Lipotoxicity; PPAR; Transcriptomics.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
References
-
- Seferovic PM, Paulus WJ. Clinical diabetic cardiomyopathy: a two-faced disease with restrictive and dilated phenotypes. Eur Heart J. 2015;36(27):1718–27, 27a-27c. - PubMed
-
- Lopaschuk GD, Folmes CD, Stanley WC. Cardiac energy metabolism in obesity. Circ Res. 2007;101(4):335–47. - PubMed
-
- Boudina S, Abel ED. Diabetic cardiomyopathy revisited. Circulation. 2007;115(25):3213–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
