

A Tgfbr1/Snai1-dependent developmental module at the core of vertebrate axial elongation
- PMID: 32597756
- PMCID: PMC7324159
- DOI: 10.7554/eLife.56615
A Tgfbr1/Snai1-dependent developmental module at the core of vertebrate axial elongation
Abstract
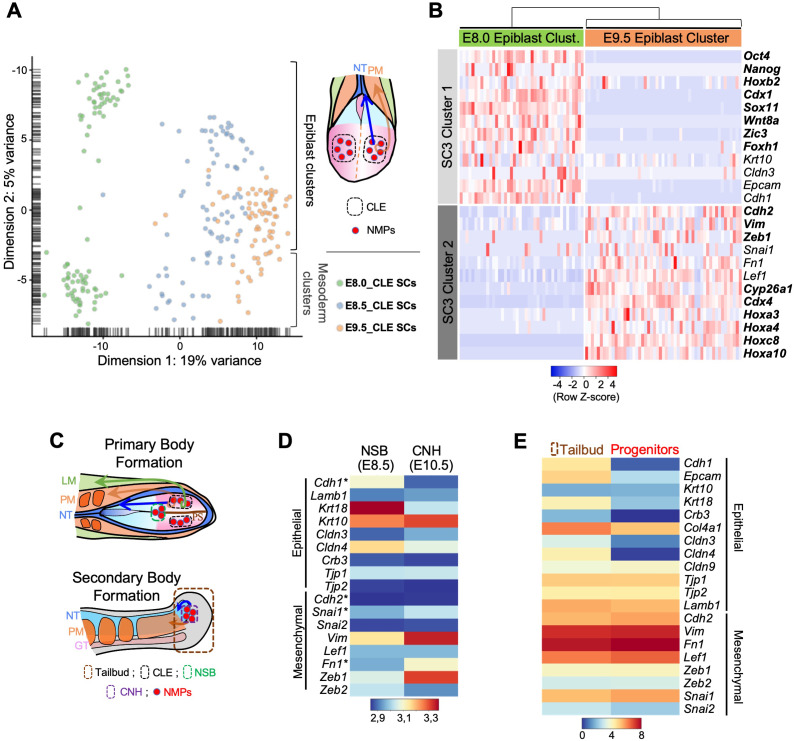
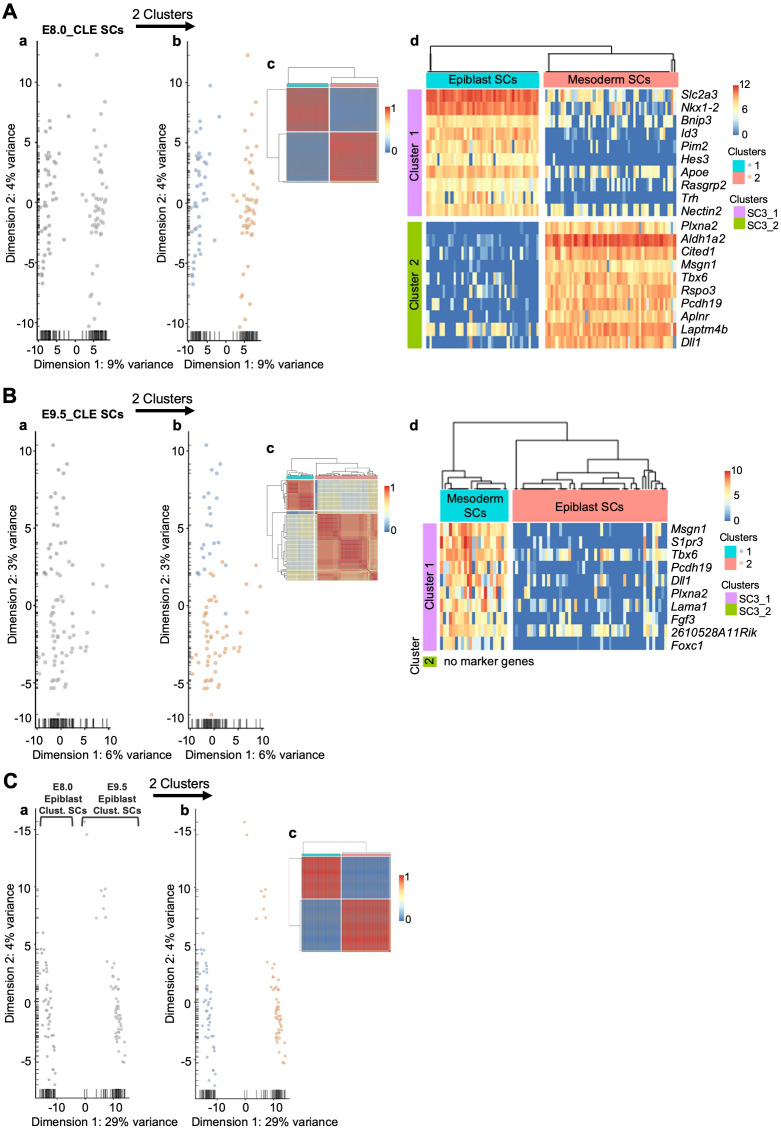
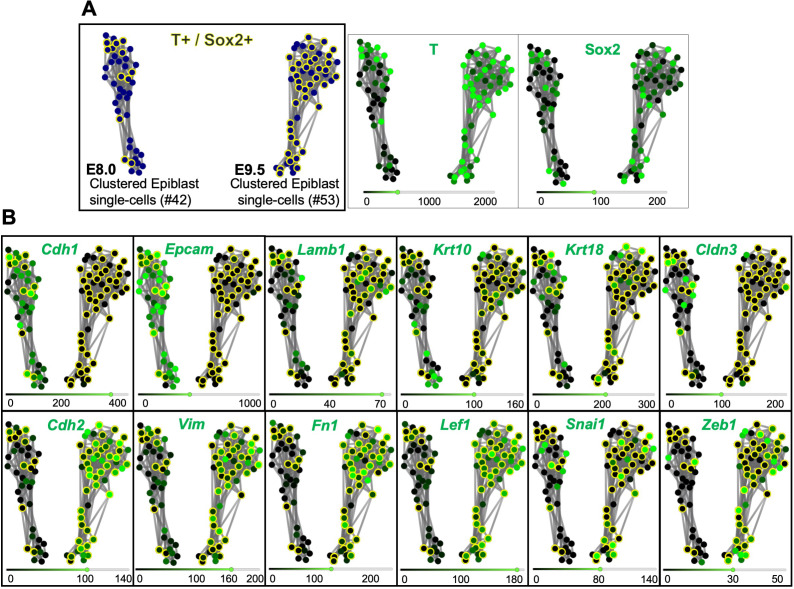
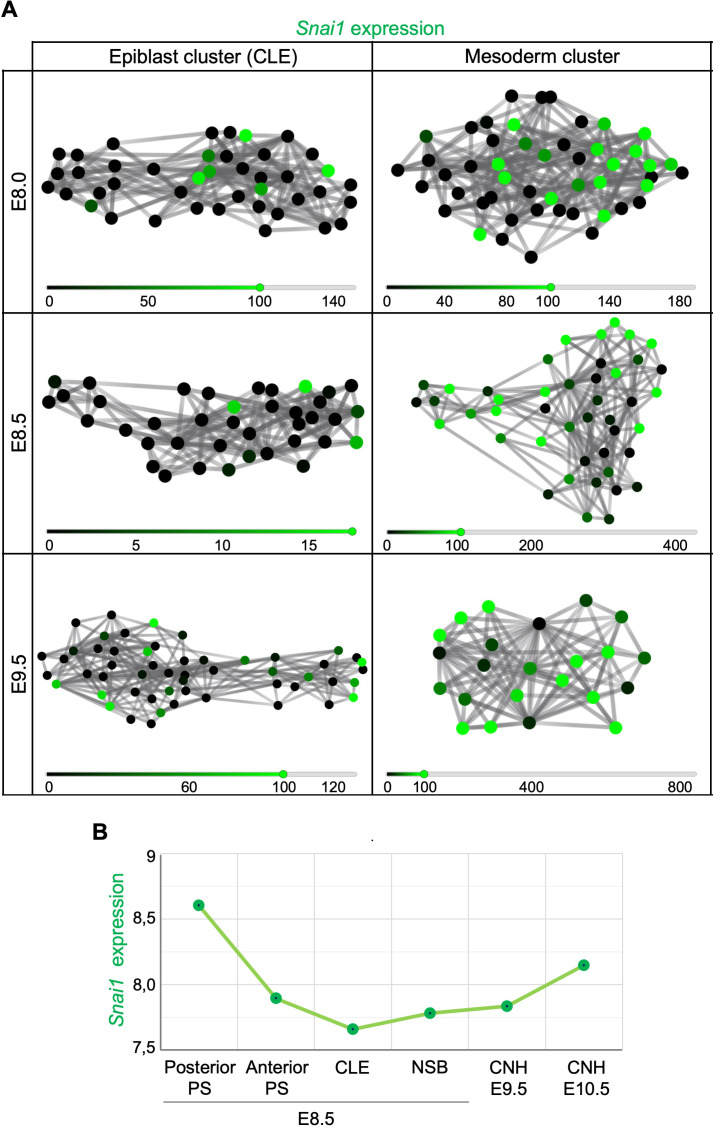
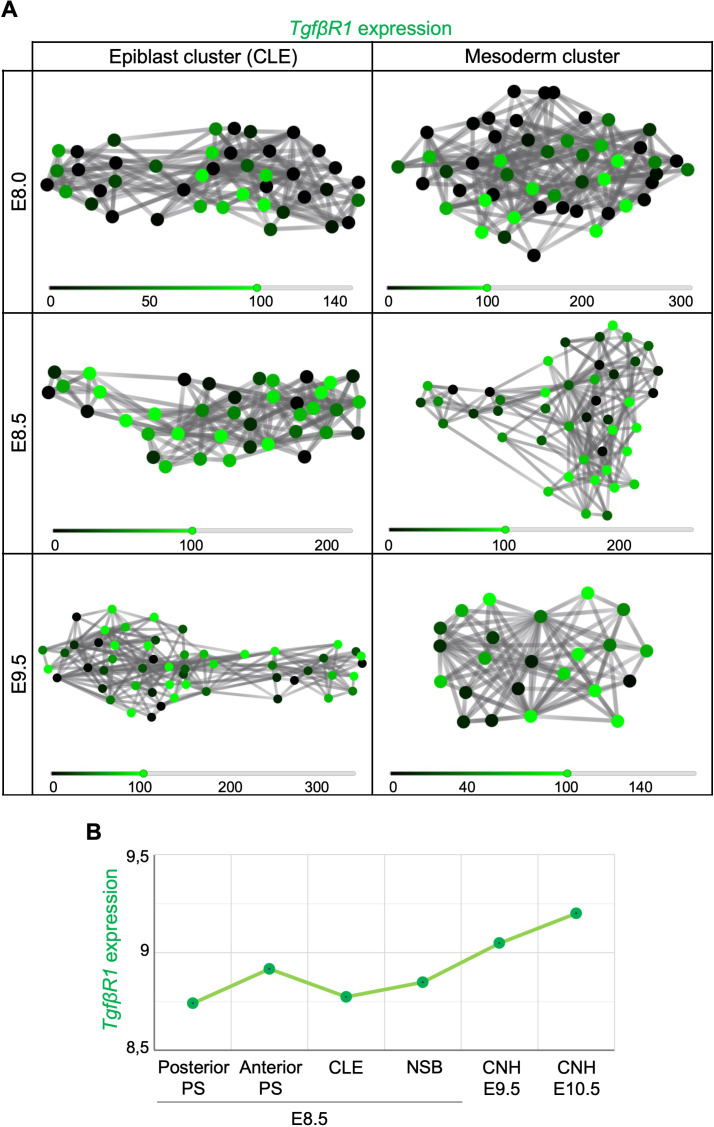
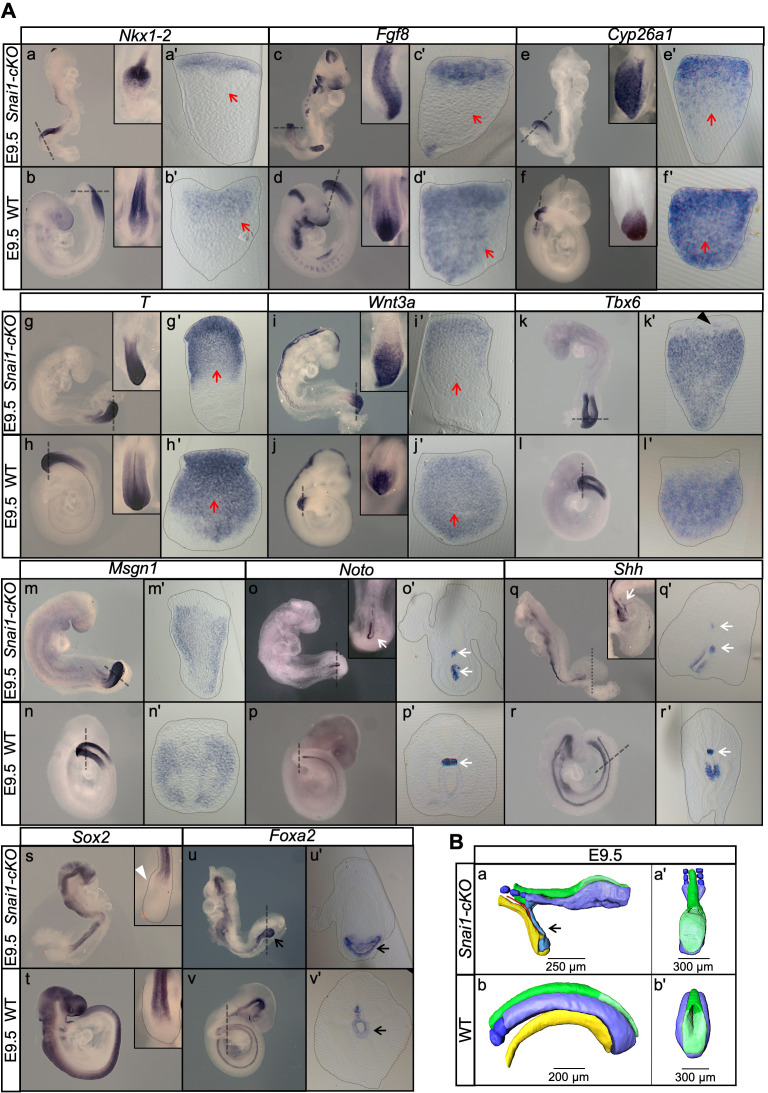
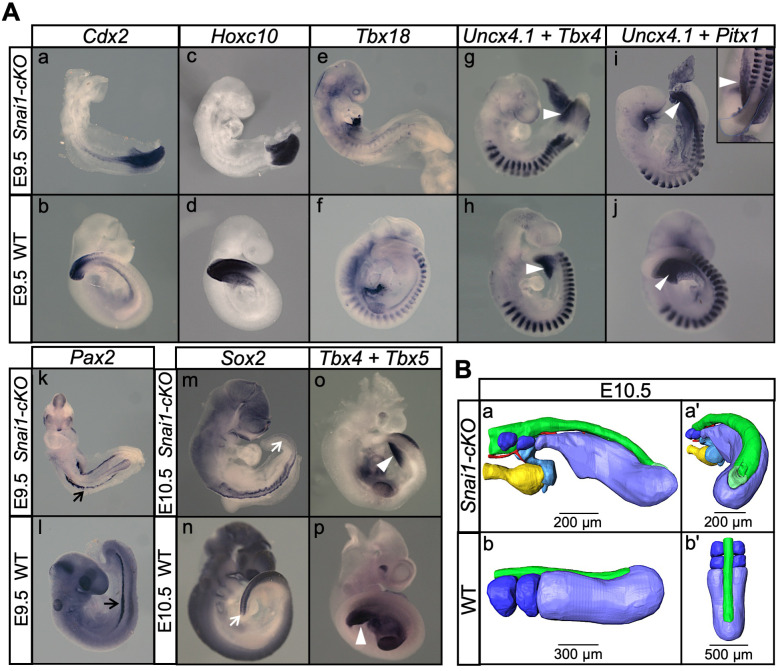
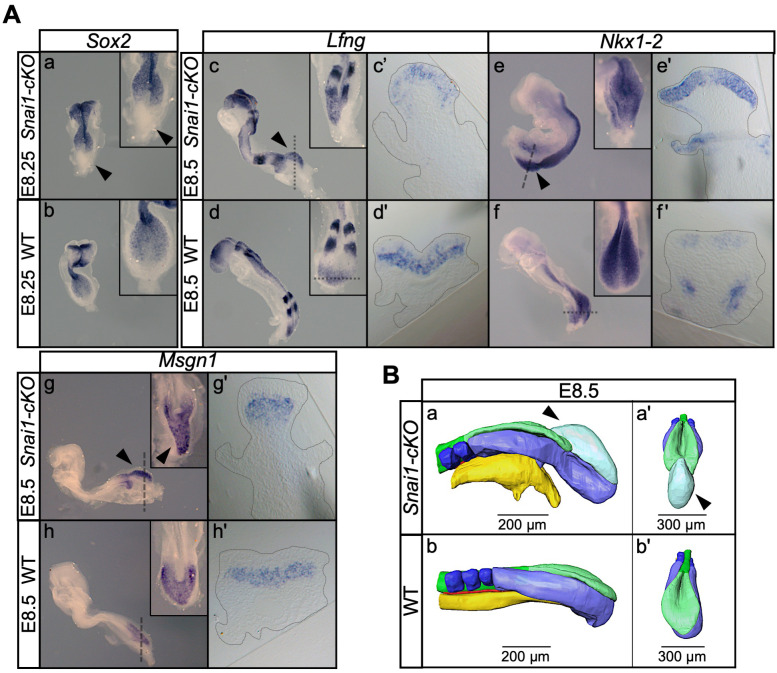
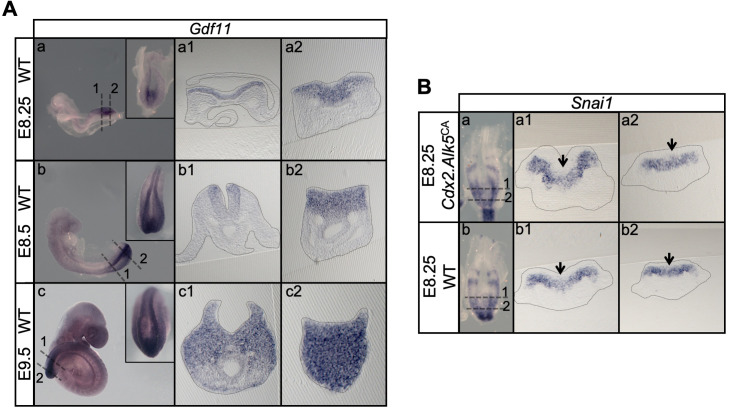
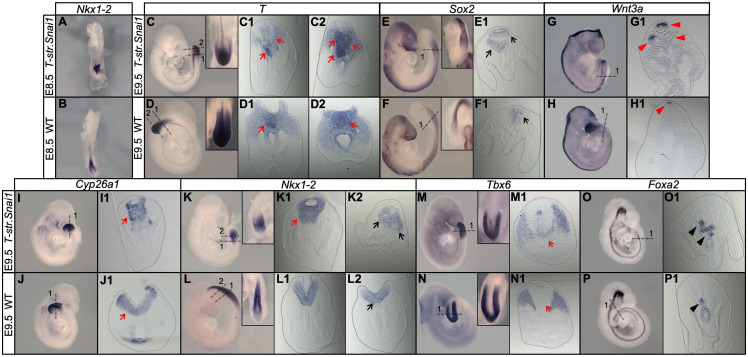
Formation of the vertebrate postcranial body axis follows two sequential but distinct phases. The first phase generates pre-sacral structures (the so-called primary body) through the activity of the primitive streak on axial progenitors within the epiblast. The embryo then switches to generate the secondary body (post-sacral structures), which depends on axial progenitors in the tail bud. Here we show that the mammalian tail bud is generated through an independent functional developmental module, concurrent but functionally different from that generating the primary body. This module is triggered by convergent Tgfbr1 and Snai1 activities that promote an incomplete epithelial to mesenchymal transition on a subset of epiblast axial progenitors. This EMT is functionally different from that coordinated by the primitive streak, as it does not lead to mesodermal differentiation but brings axial progenitors into a transitory state, keeping their progenitor activity to drive further axial body extension.
Keywords: EMT; axial elongation; axial progenitors; developmental biology; mouse; snai1; tail bud; tgfbr1.
© 2020, Dias et al.
Conflict of interest statement
AD, AL, FW, AN, AB, DS, GM, VW, MM No competing interests declared
Figures
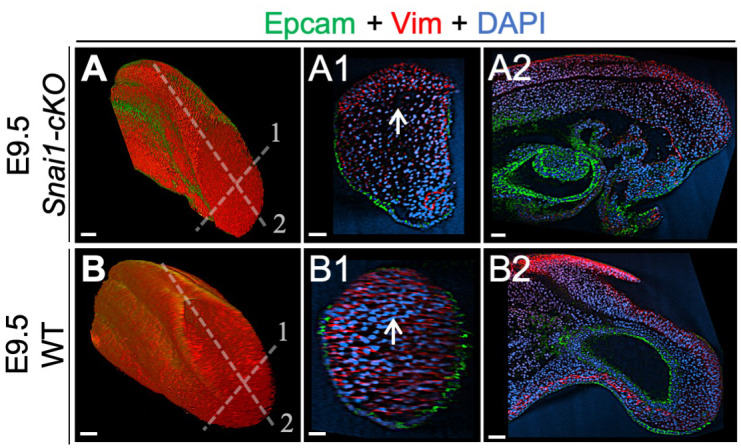
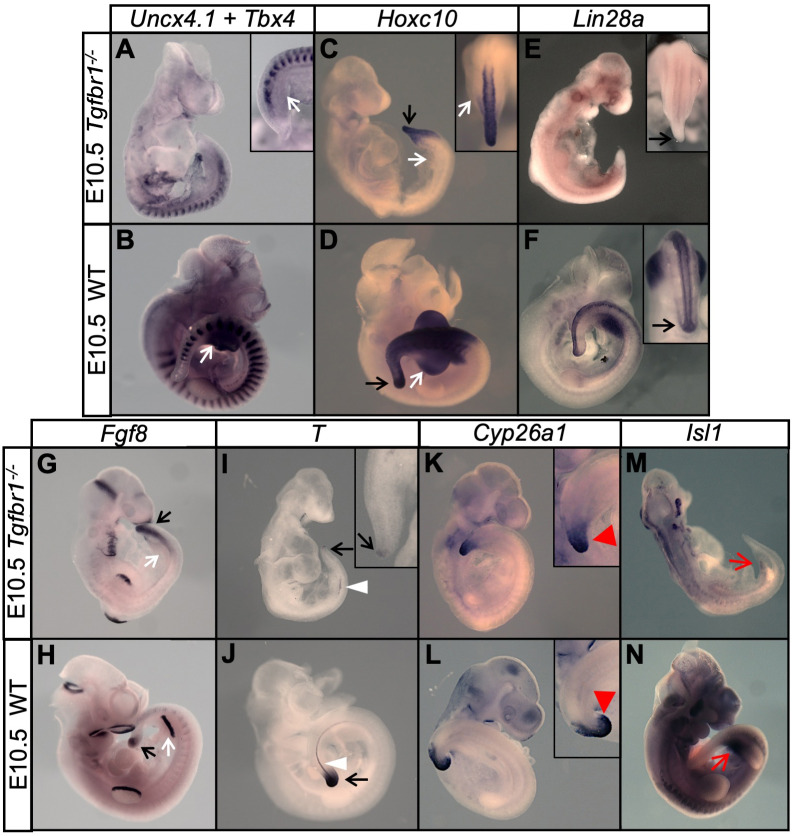
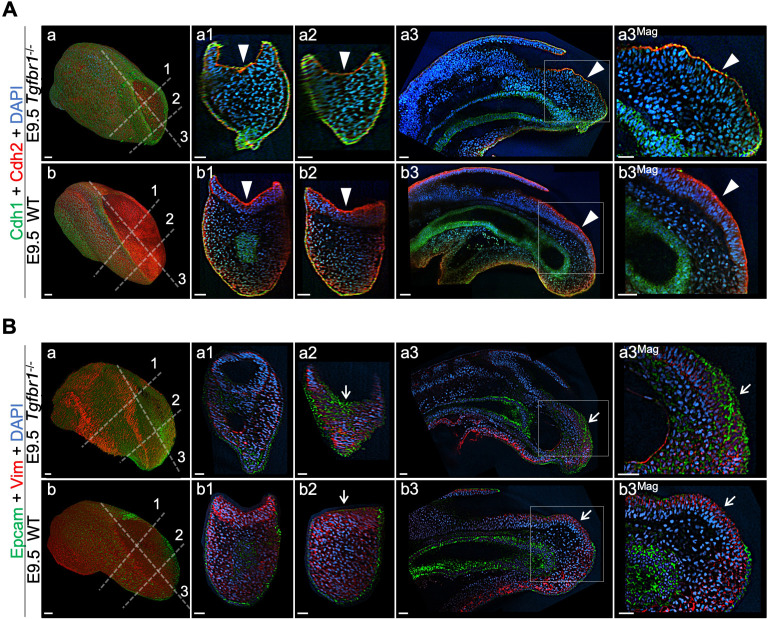
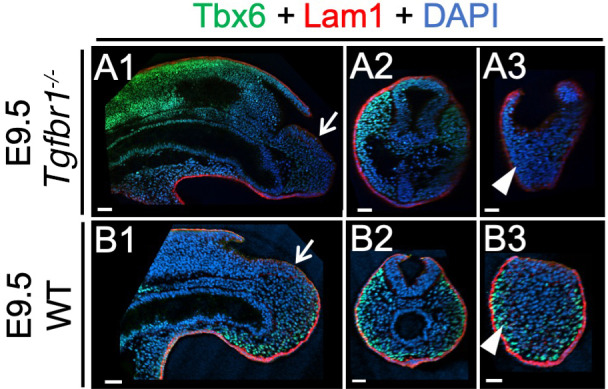
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- SRA/SRX4968732
- SRA/SRX4968731
- SRA/SRX5532193
- SRA/SRX5532192
Grants and funding
- MR/S008799/1/MRC_/Medical Research Council/United Kingdom
- PTDC/BEX-BID/0899/2014/Fundação para a Ciência e a Tecnologia/International
- LISBOA-01-0145-FEDER-030254/Fundação para a Ciência e a Tecnologia/International
- DEV-170806/Company of Biologists/International
- LISBOA-01-0145-FEDER-022170/Fundação para a Ciência e a Tecnologia/International
- MR/K011200/1/MRC_/Medical Research Council/United Kingdom
- SCML-MC-60-2014/Santa Casa da Misericórdia/International
- MR/K017047/1/MRC_/Medical Research Council/United Kingdom
- PD/BD/128437/2017/Fundação para a Ciência e a Tecnologia/International
- PD/BD/128426/2017/Fundação para a Ciência e a Tecnologia/International
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
