

The methyltransferase TrmA facilitates tRNA folding through interaction with its RNA-binding domain
- PMID: 32597953
- PMCID: PMC7641329
- DOI: 10.1093/nar/gkaa548
The methyltransferase TrmA facilitates tRNA folding through interaction with its RNA-binding domain
Abstract
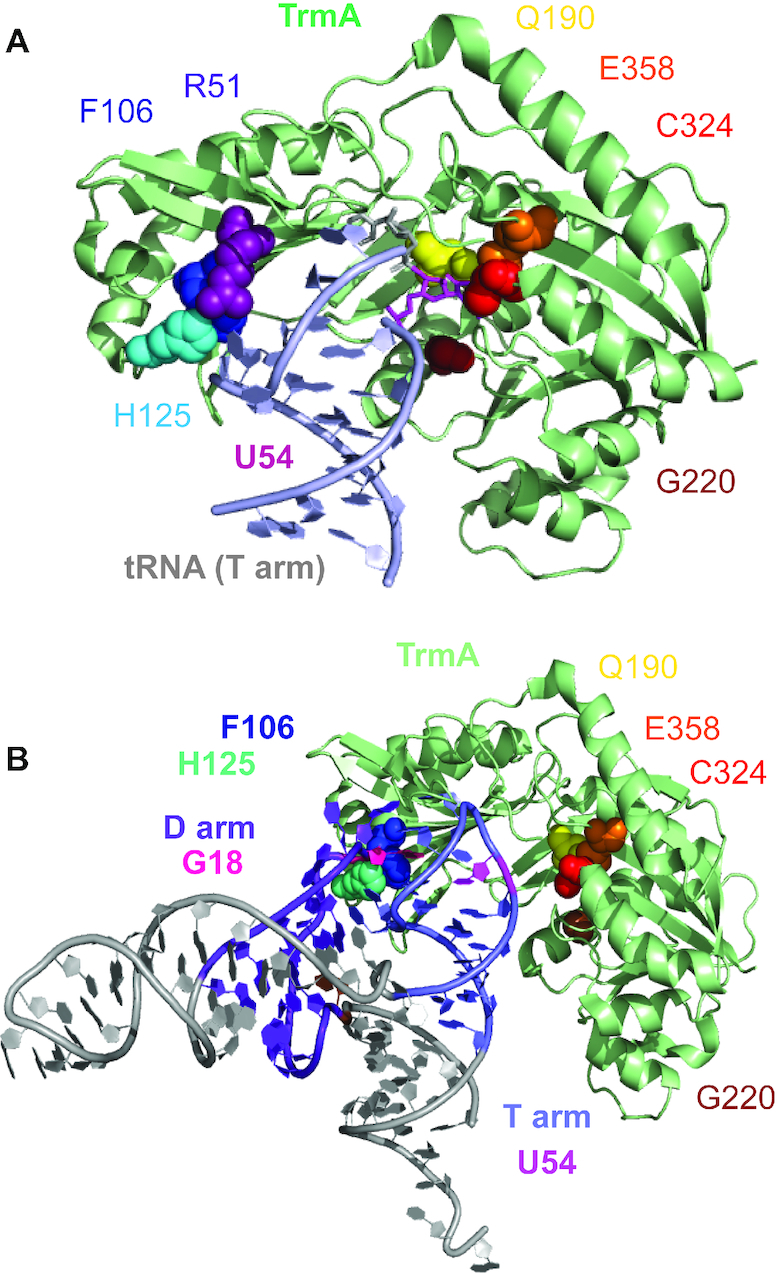
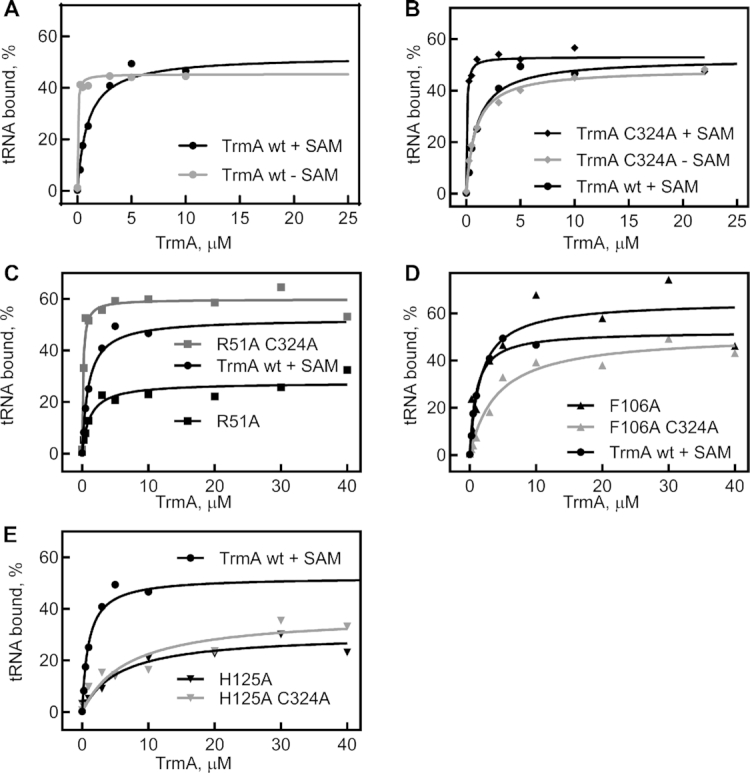
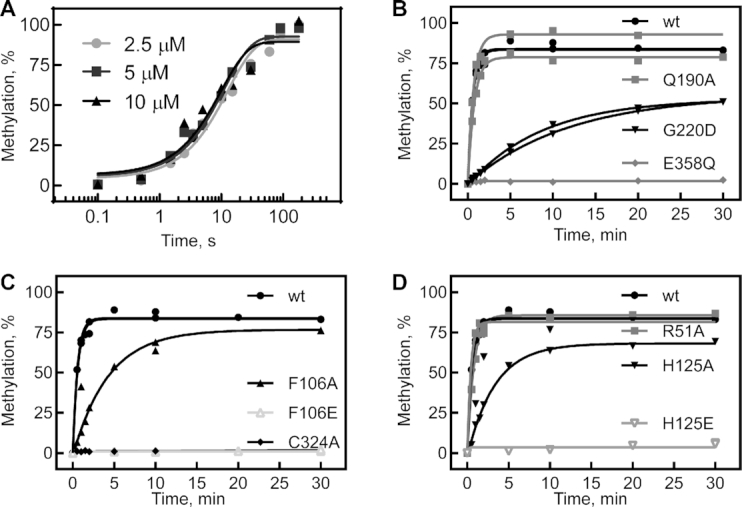
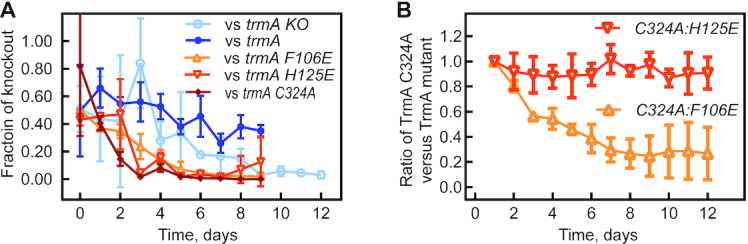
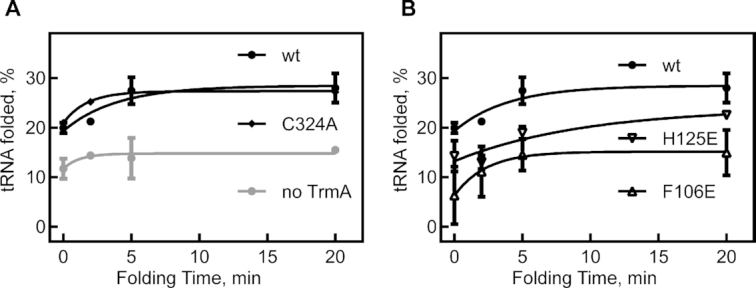
tRNAs are the most highly modified RNAs in all cells, and formation of 5-methyluridine (m5U) at position 54 in the T arm is a common RNA modification found in all tRNAs. The m5U modification is generated by the methyltransferase TrmA. Here, we test and prove the hypothesis that Escherichia coli TrmA has dual functions, acting both as a methyltransferase and as a tRNA chaperone. We identify two conserved residues, F106 and H125, in the RNA-binding domain of TrmA, which interact with the tRNA elbow and are critical for tRNA binding. Co-culture competition assays reveal that the catalytic activity of TrmA is important for cellular fitness, and that substitutions of F106 or H125 impair cellular fitness. We directly show that TrmA enhances tRNA folding in vitro independent of its catalytic activity. In conclusion, our study suggests that F106 and H125 in the RNA-binding domain of TrmA act as a wedge disrupting tertiary interactions between tRNA's D arm and T arm; this tRNA unfolding is the mechanistic basis for TrmA's tRNA chaperone activity. TrmA is the second tRNA modifying enzyme next to the pseudouridine synthase TruB shown to act as a tRNA chaperone supporting a functional link between RNA modification and folding.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Gutgsell N., Englund N., Niu L., Kaya Y., Lane B.G., Ofengand J.. Deletion of the Escherichia coli pseudouridine synthase gene truB blocks formation of pseudouridine 55 in tRNA in vivo, does not affect exponential growth, but confers a strong selective disadvantage in competition with wild-type cells. RNA. 2000; 6:1870–1881. - PMC - PubMed
-
- Ishitani R., Yokoyama S., Nureki O.. Structure, dynamics, and function of RNA modification enzymes. Curr. Opin. Struct. Biol. 2008; 18:330–339. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
