

Developmental onset distinguishes three types of spontaneous recognition memory in mice
- PMID: 32606443
- PMCID: PMC7326931
- DOI: 10.1038/s41598-020-67619-w
Developmental onset distinguishes three types of spontaneous recognition memory in mice
Abstract
Spontaneous recognition memory tasks build on an animal's natural preference for novelty to assess the what, where and when components of episodic memory. Their simplicity, ethological relevance and cross-species adaptability make them extremely useful to study the physiology and pathology of memory. Recognition memory deficits are common in rodent models of neurodevelopmental disorders, and yet very little is known about the expression of spontaneous recognition memory in young rodents. This is exacerbated by the paucity of data on the developmental onset of recognition memory in mice, a major animal model of disease. To address this, we characterized the ontogeny of three types of spontaneous recognition memory in mice: object location, novel object recognition and temporal order recognition. We found that object location is the first to emerge, at postnatal day (P)21. This was followed by novel object recognition (24 h delay), at P25. Temporal order recognition was the last to emerge, at P28. Elucidating the developmental expression of recognition memory in mice is critical to improving our understanding of the ontogeny of episodic memory, and establishes a necessary blueprint to apply these tasks to probe cognitive deficits at clinically relevant time points in animal models of developmental disorders.
Conflict of interest statement
The authors declare no competing interests.
Figures
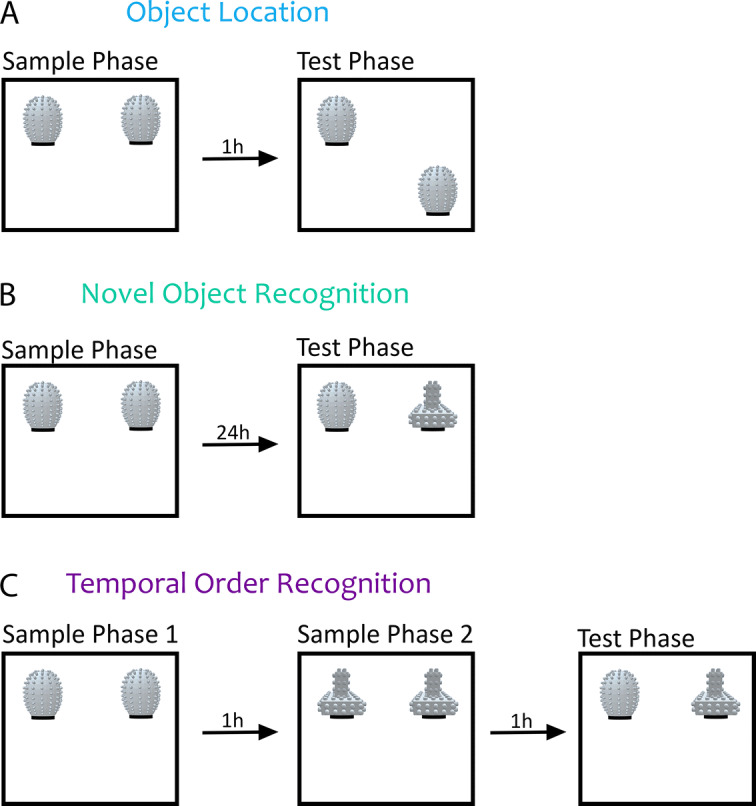
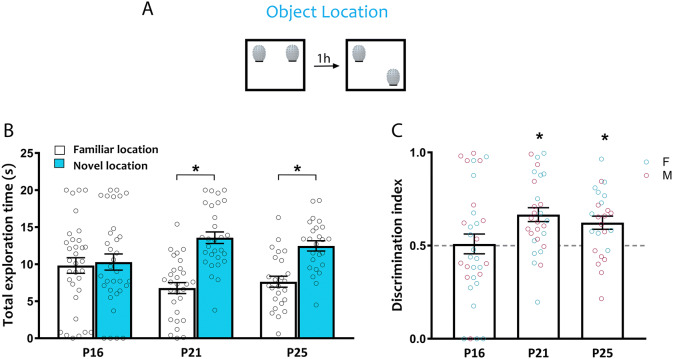
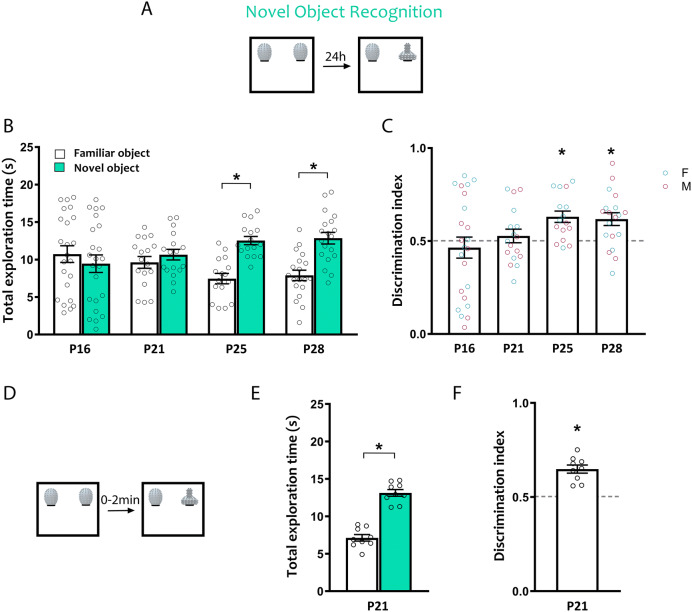

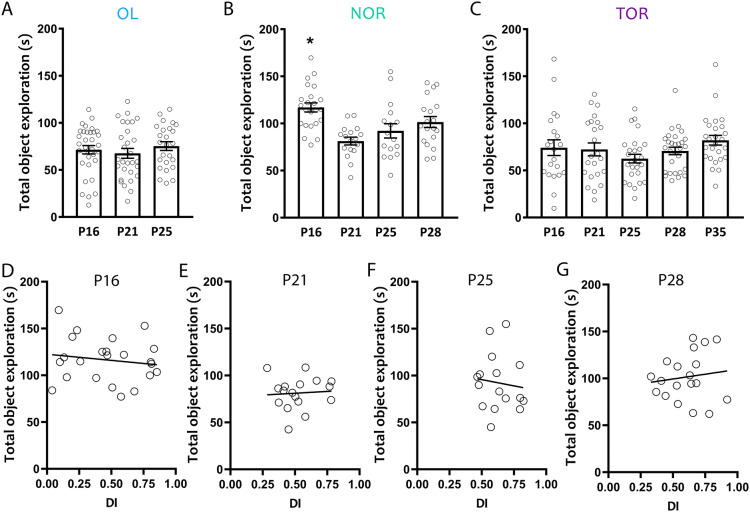
References
-
- Winters BD, Saksida LM, Bussey TJ. Object recognition memory: Neurobiological mechanisms of encoding, consolidation and retrieval. Neurosci. Biobehav. Rev. 2008;32:1055–1070. - PubMed
-
- Ennaceur A, Delacour J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988;31:47–59. - PubMed
-
- Bevins RA, Besheer J. Object recognition in rats and mice: A one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nat. Protoc. 2006;1:1306–1311. - PubMed
-
- Dere E, Huston JP, De Souza Silva MA. The pharmacology, neuroanatomy and neurogenetics of one-trial object recognition in rodents. Neurosci. Biobehav. Rev. 2007;31:673–704. - PubMed
-
- Alvarado MC, Bachevalier J. Revisiting the Maturation of medial temporal lobe memory functions in primates. Learn. Mem. 2000;7:244–256. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
