

Diversification in evolutionary arenas-Assessment and synthesis
- PMID: 32607221
- PMCID: PMC7319112
- DOI: 10.1002/ece3.6313
Diversification in evolutionary arenas-Assessment and synthesis
Abstract
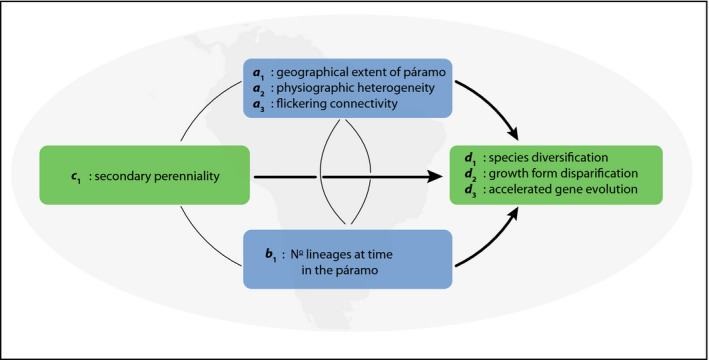
Understanding how and why rates of evolutionary diversification vary is a key issue in evolutionary biology, ecology, and biogeography. Evolutionary rates are the net result of interacting processes summarized under concepts such as adaptive radiation and evolutionary stasis. Here, we review the central concepts in the evolutionary diversification literature and synthesize these into a simple, general framework for studying rates of diversification and quantifying their underlying dynamics, which can be applied across clades and regions, and across spatial and temporal scales. Our framework describes the diversification rate (d) as a function of the abiotic environment (a), the biotic environment (b), and clade-specific phenotypes or traits (c); thus, d ~ a,b,c. We refer to the four components (a-d) and their interactions collectively as the "Evolutionary Arena." We outline analytical approaches to this framework and present a case study on conifers, for which we parameterize the general model. We also discuss three conceptual examples: the Lupinus radiation in the Andes in the context of emerging ecological opportunity and fluctuating connectivity due to climatic oscillations; oceanic island radiations in the context of island formation and erosion; and biotically driven radiations of the Mediterranean orchid genus Ophrys. The results of the conifer case study are consistent with the long-standing scenario that low competition and high rates of niche evolution promote diversification. The conceptual examples illustrate how using the synthetic Evolutionary Arena framework helps to identify and structure future directions for research on evolutionary radiations. In this way, the Evolutionary Arena framework promotes a more general understanding of variation in evolutionary rates by making quantitative results comparable between case studies, thereby allowing new syntheses of evolutionary and ecological processes to emerge.
Keywords: adaptive radiation; conifer phylogeny; macroevolutionary theory; phylogenetic comparative methods; species diversification; trait disparification.
© 2020 The Authors. Ecology and Evolution published by John Wiley & Sons Ltd.
Conflict of interest statement
None declared.
Figures





References
-
- Alfaro, M. E. , Santini, F. , Brock, C. , Alamillo, H. , Dornburg, A. , Rabosky, D. L. , … Harmon, L. J. (2009). Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. Proceedings of the National Academy of Sciences of the United States of America, 106(32), 13410–13414. 10.1073/pnas.0811087106 - DOI - PMC - PubMed
-
- Arakaki, M. , Christin, P.‐A. , Nyffeler, R. , Lendel, A. , Eggli, U. , Ogburn, R. M. , … Edwards, E. J. (2011). Contemporaneous and recent radiations of the world's major succulent plant lineages. Proceedings of the National Academy of Sciences of the United States of America, 108(20), 8379–8384. 10.1073/pnas.1100628108 - DOI - PMC - PubMed
Publication types
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
