

Transcriptional and Functional Analysis of CD1c+ Human Dendritic Cells Identifies a CD163+ Subset Priming CD8+CD103+ T Cells
- PMID: 32610077
- PMCID: PMC7445430
- DOI: 10.1016/j.immuni.2020.06.002
Transcriptional and Functional Analysis of CD1c+ Human Dendritic Cells Identifies a CD163+ Subset Priming CD8+CD103+ T Cells
Abstract
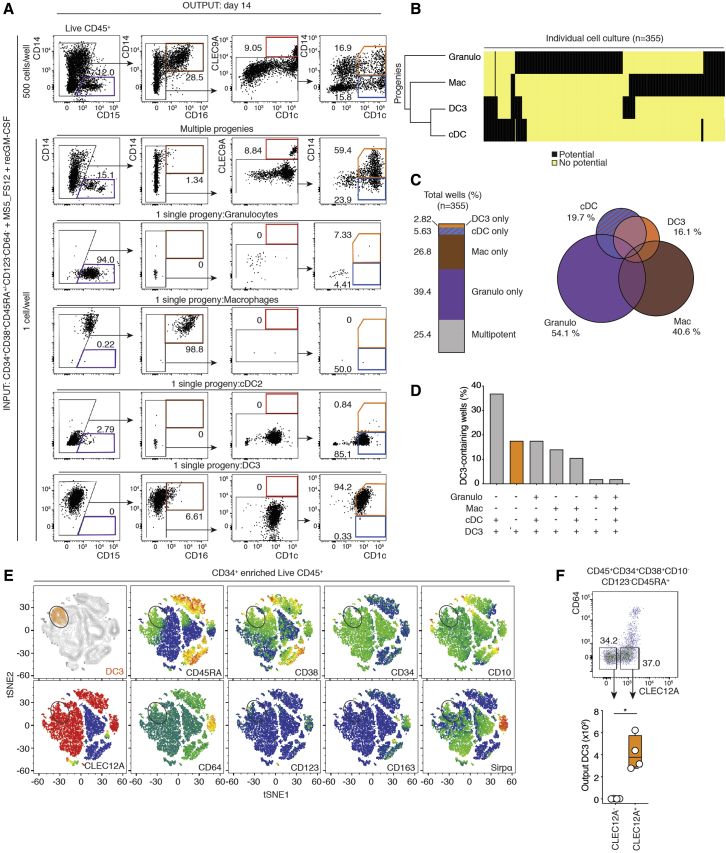
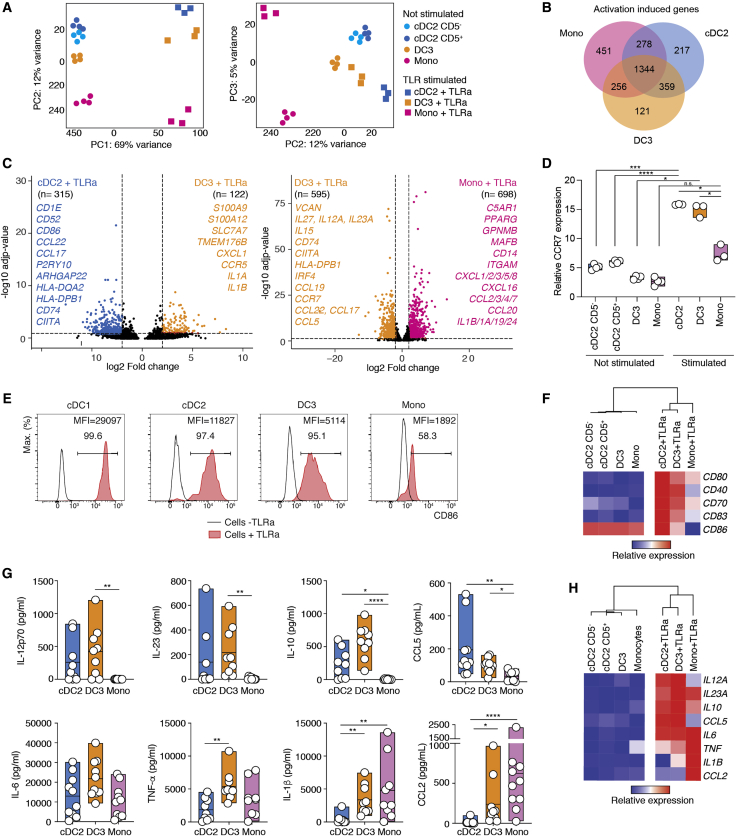
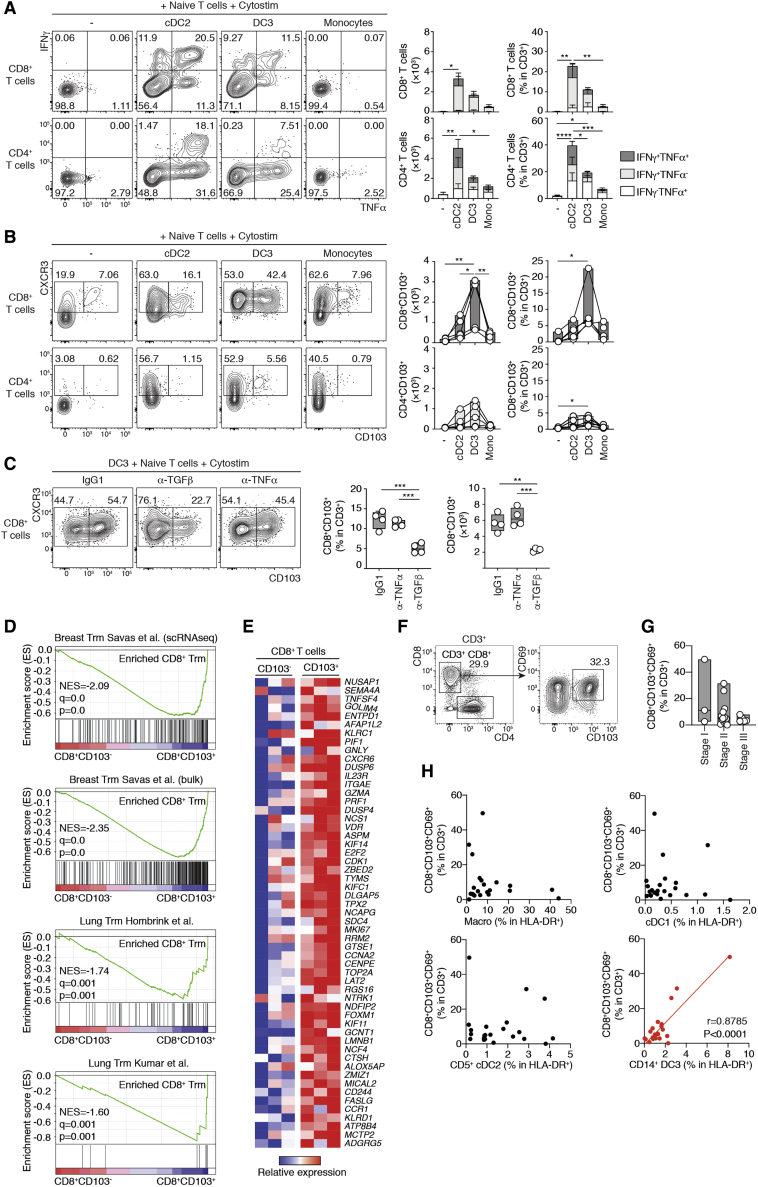
Dendritic cells (DCs) are antigen-presenting cells controlling T cell activation. In humans, the diversity, ontogeny, and functional capabilities of DC subsets are not fully understood. Here, we identified circulating CD88-CD1c+CD163+ DCs (called DC3s) as immediate precursors of inflammatory CD88-CD14+CD1c+CD163+FcεRI+ DCs. DC3s develop via a specific pathway activated by GM-CSF, independent of cDC-restricted (CDP) and monocyte-restricted (cMoP) progenitors. Like classical DCs but unlike monocytes, DC3s drove activation of naive T cells. In vitro, DC3s displayed a distinctive ability to prime CD8+ T cells expressing a tissue homing signature and the epithelial homing alpha-E integrin (CD103) through transforming growth factor β (TGF-β) signaling. In vivo, DC3s infiltrated luminal breast cancer primary tumors, and DC3 infiltration correlated positively with CD8+CD103+CD69+ tissue-resident memory T cells. Together, these findings define DC3s as a lineage of inflammatory DCs endowed with a strong potential to regulate tumor immunity.
Keywords: DC progenitors; DC3s; T(RM); cDC2s; conventional DCs; inflammatory DCs; monocytes; mononuclear phagocytes.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures





Comment in
-
The More, the Merrier: DC3s Join the Human Dendritic Cell Family.Immunity. 2020 Aug 18;53(2):233-235. doi: 10.1016/j.immuni.2020.07.014. Immunity. 2020. PMID: 32814019
References
-
- Acosta-Rodriguez E.V., Napolitani G., Lanzavecchia A., Sallusto F. Interleukins 1beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat. Immunol. 2007;8:942–949. - PubMed
-
- Bakdash G., Buschow S.I., Gorris M.A., Halilovic A., Hato S.V., Sköld A.E., Schreibelt G., Sittig S.P., Torensma R., Duiveman-de Boer T. Expansion of a BDCA1+CD14+ Myeloid Cell Population in Melanoma Patients May Attenuate the Efficacy of Dendritic Cell Vaccines. Cancer Res. 2016;76:4332–4346. - PubMed
-
- Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
