

Ribosome Collisions Trigger General Stress Responses to Regulate Cell Fate
- PMID: 32610081
- PMCID: PMC7384957
- DOI: 10.1016/j.cell.2020.06.006
Ribosome Collisions Trigger General Stress Responses to Regulate Cell Fate
Abstract
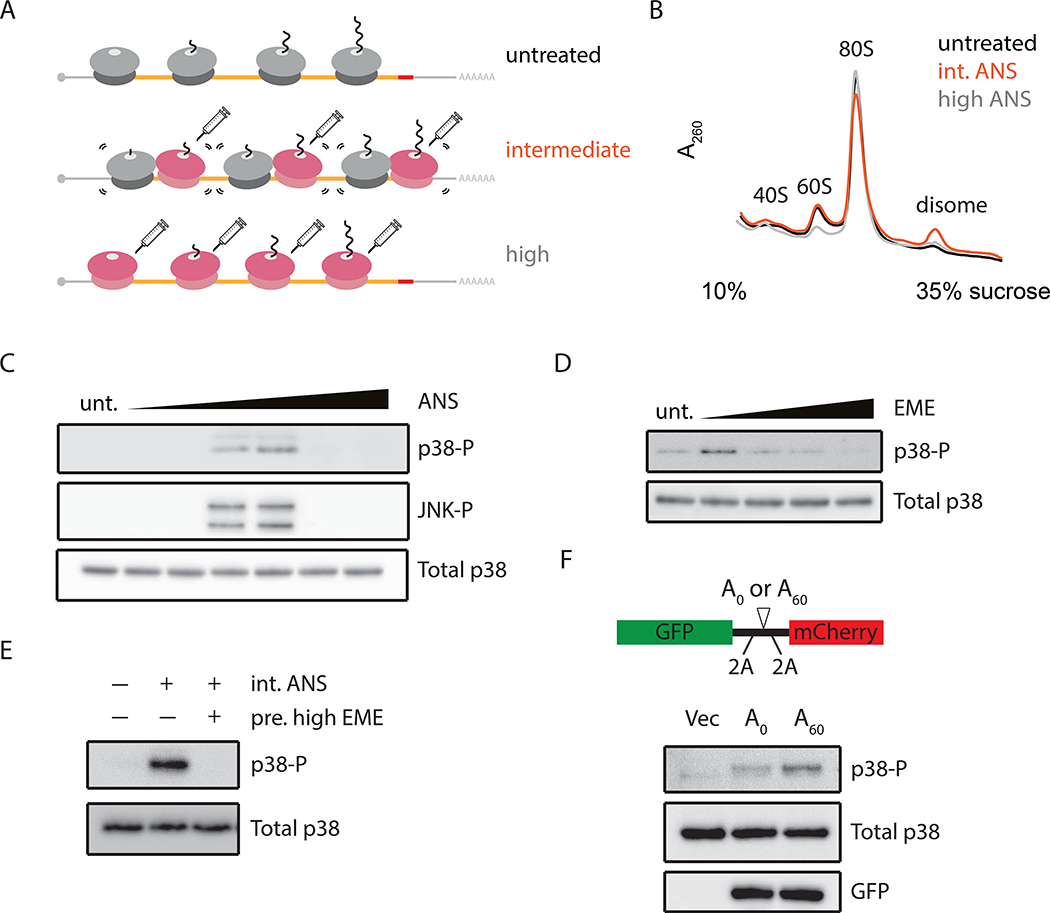
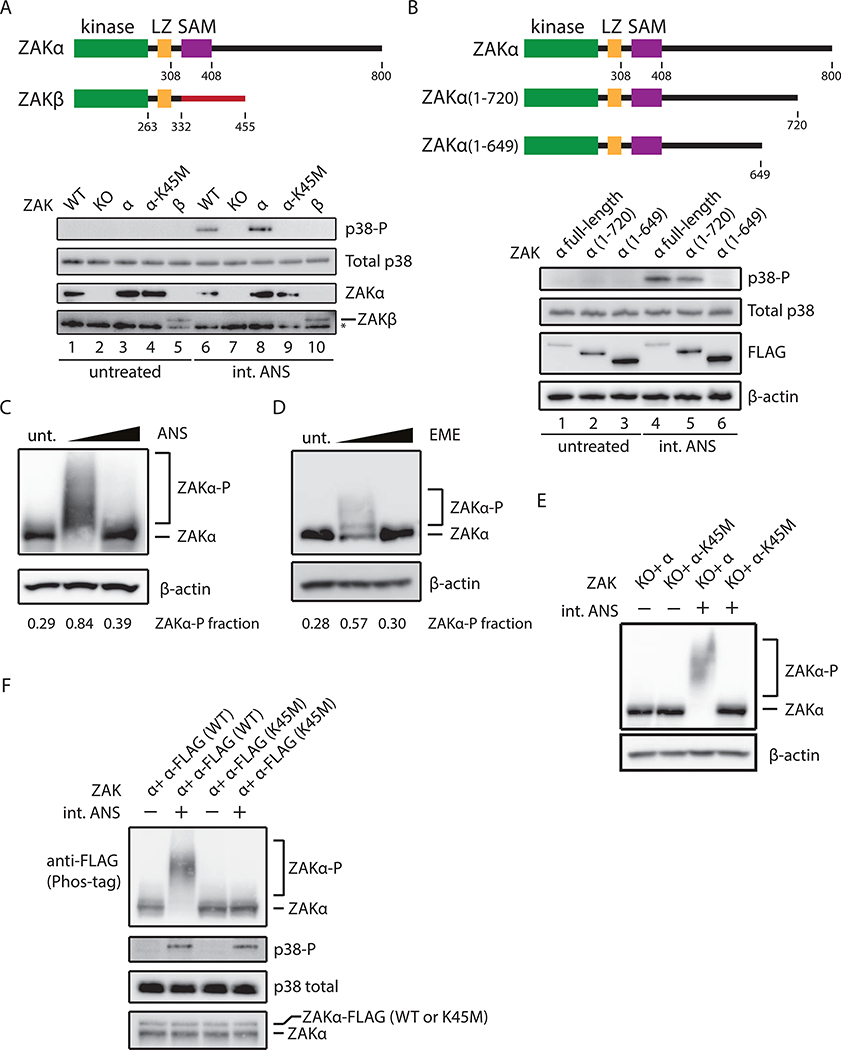
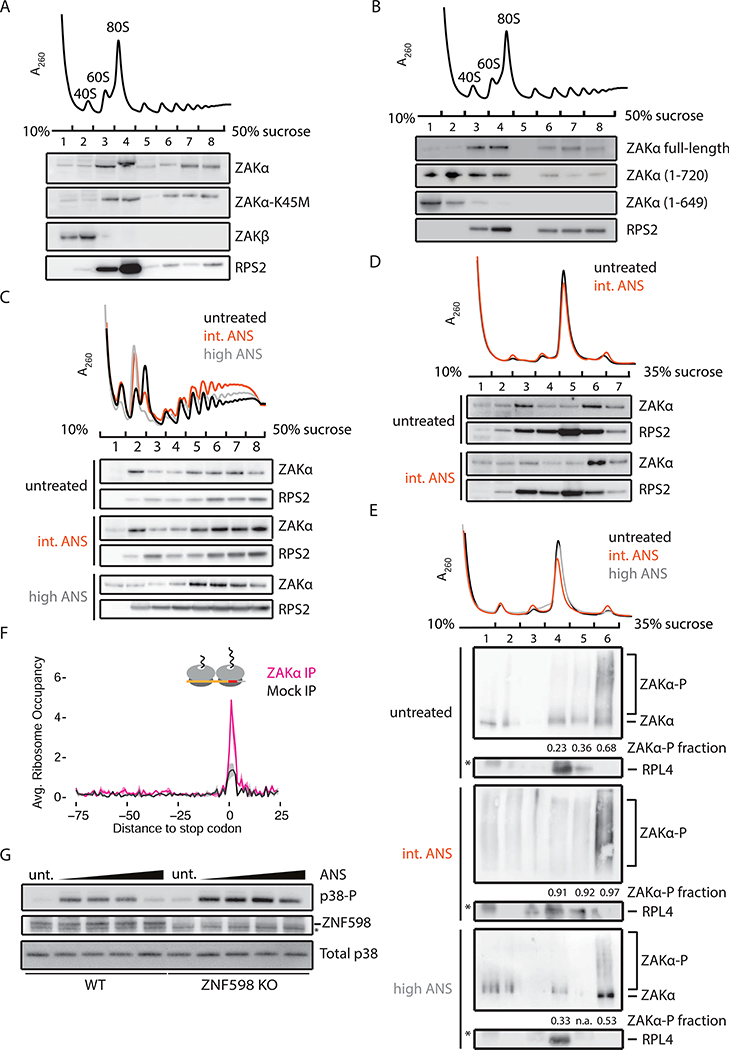
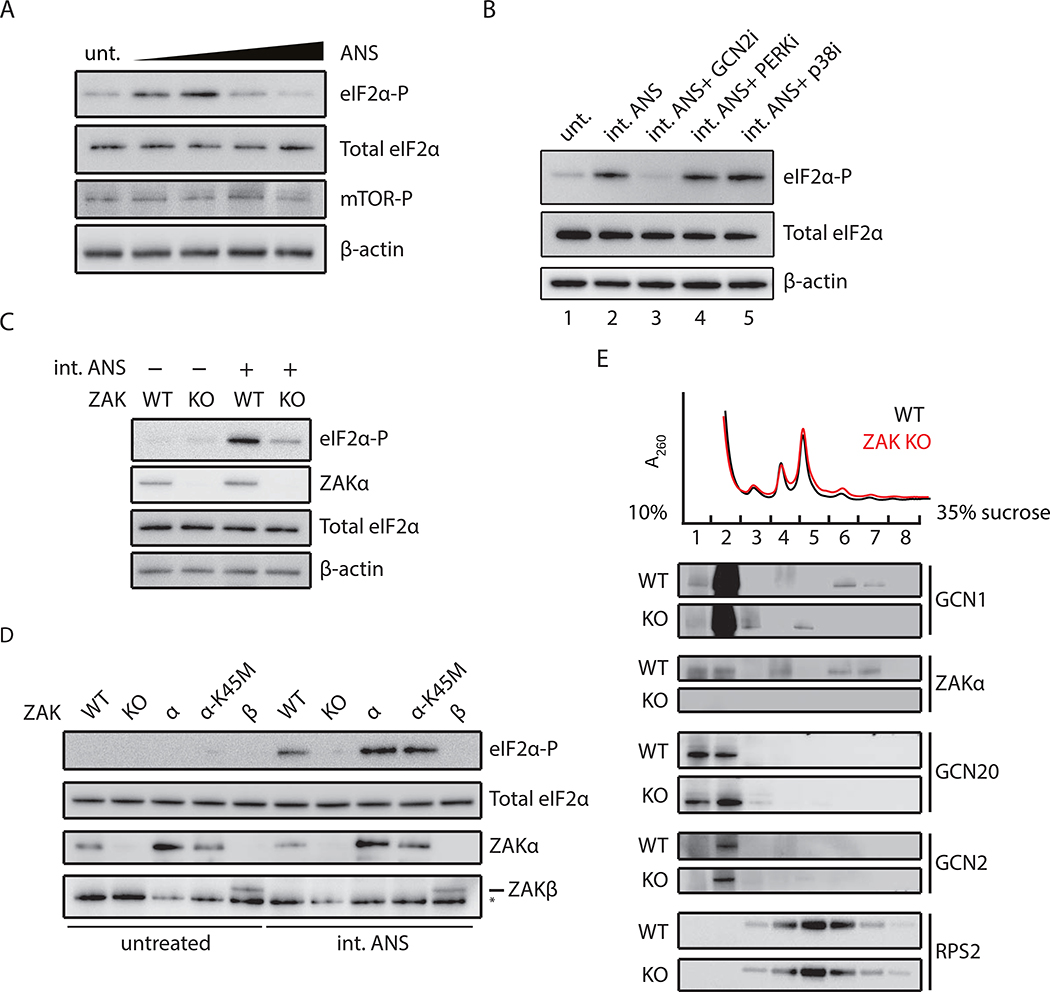
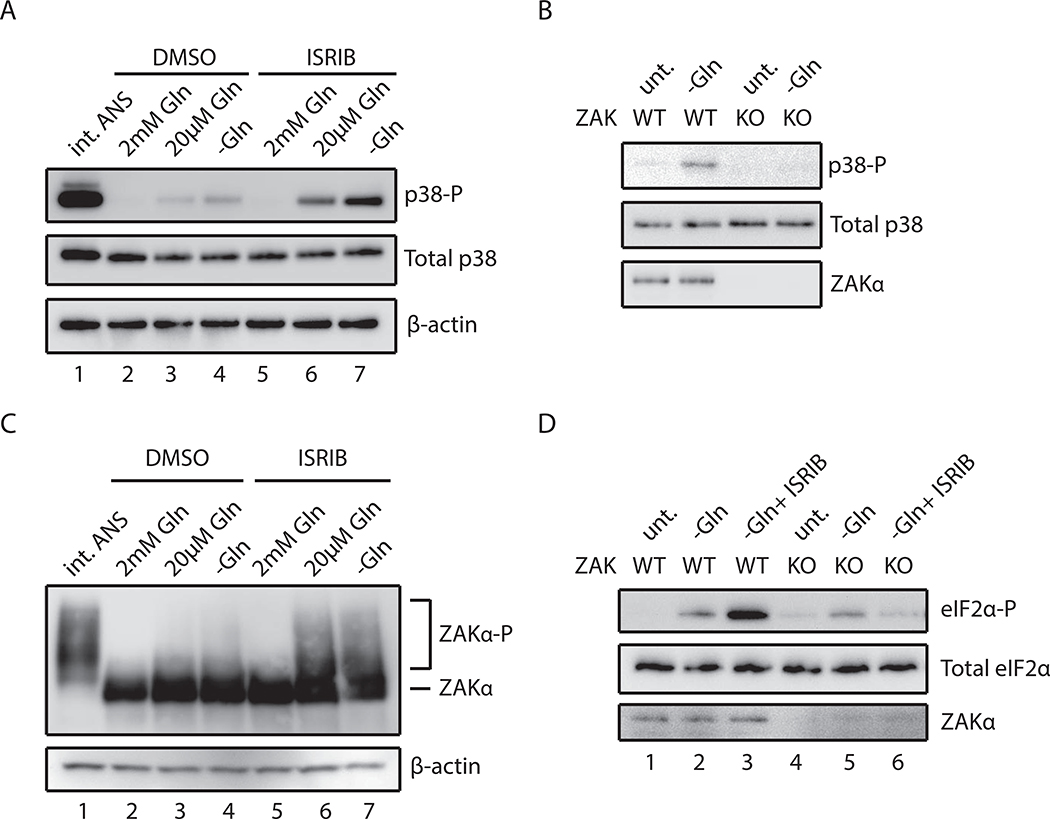
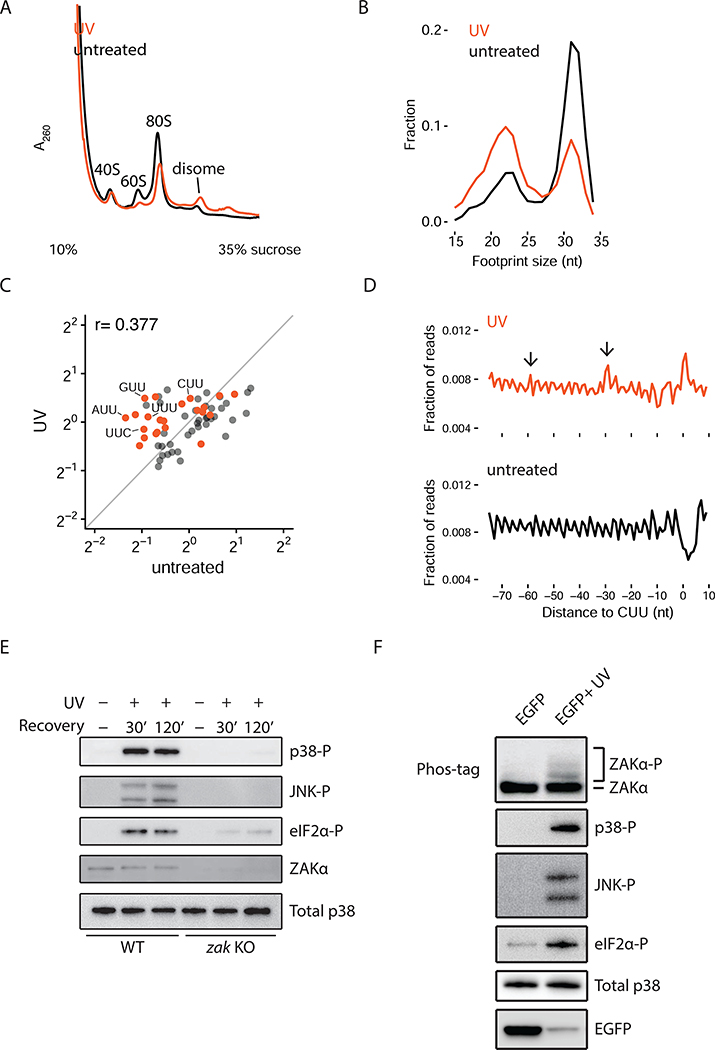
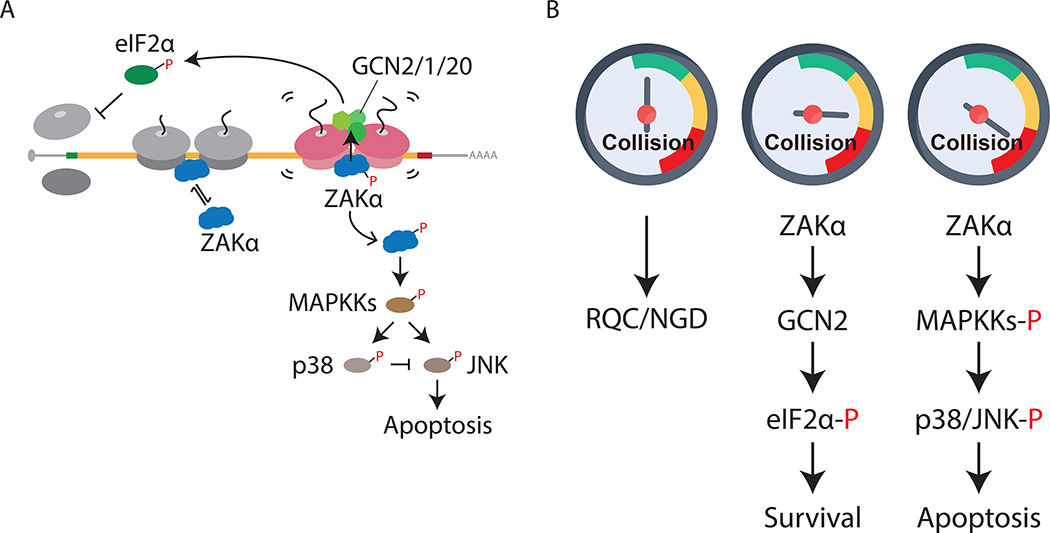
Problems arising during translation of mRNAs lead to ribosome stalling and collisions that trigger a series of quality control events. However, the global cellular response to ribosome collisions has not been explored. Here, we uncover a function for ribosome collisions in signal transduction. Using translation elongation inhibitors and general cellular stress conditions, including amino acid starvation and UV irradiation, we show that ribosome collisions activate the stress-activated protein kinase (SAPK) and GCN2-mediated stress response pathways. We show that the MAPKKK ZAK functions as the sentinel for ribosome collisions and is required for immediate early activation of both SAPK (p38/JNK) and GCN2 signaling pathways. Selective ribosome profiling and biochemistry demonstrate that although ZAK generally associates with elongating ribosomes on polysomal mRNAs, it specifically auto-phosphorylates on the minimal unit of colliding ribosomes, the disome. Together, these results provide molecular insights into how perturbation of translational homeostasis regulates cell fate.
Keywords: SAPK; UV radiation; ZAK; amino acid starvation; integrated stress response; ribosome collisions.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Darling NJ, and Cook SJ (2014). The role of MAPK signalling pathways in the response to endoplasmic reticulum stress. Biochim. Biophys. Acta - Mol. Cell Res. 1843, 2150–2163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
