

Genetically and Antigenically Divergent Influenza A(H9N2) Viruses Exhibit Differential Replication and Transmission Phenotypes in Mammalian Models
- PMID: 32611751
- PMCID: PMC7431790
- DOI: 10.1128/JVI.00451-20
Genetically and Antigenically Divergent Influenza A(H9N2) Viruses Exhibit Differential Replication and Transmission Phenotypes in Mammalian Models
Abstract
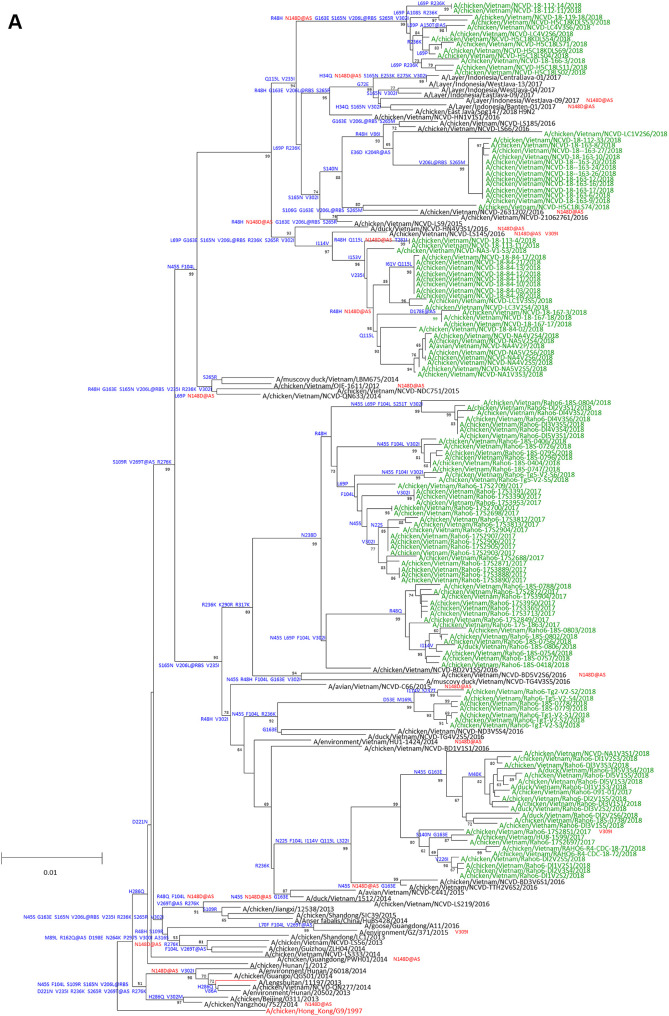
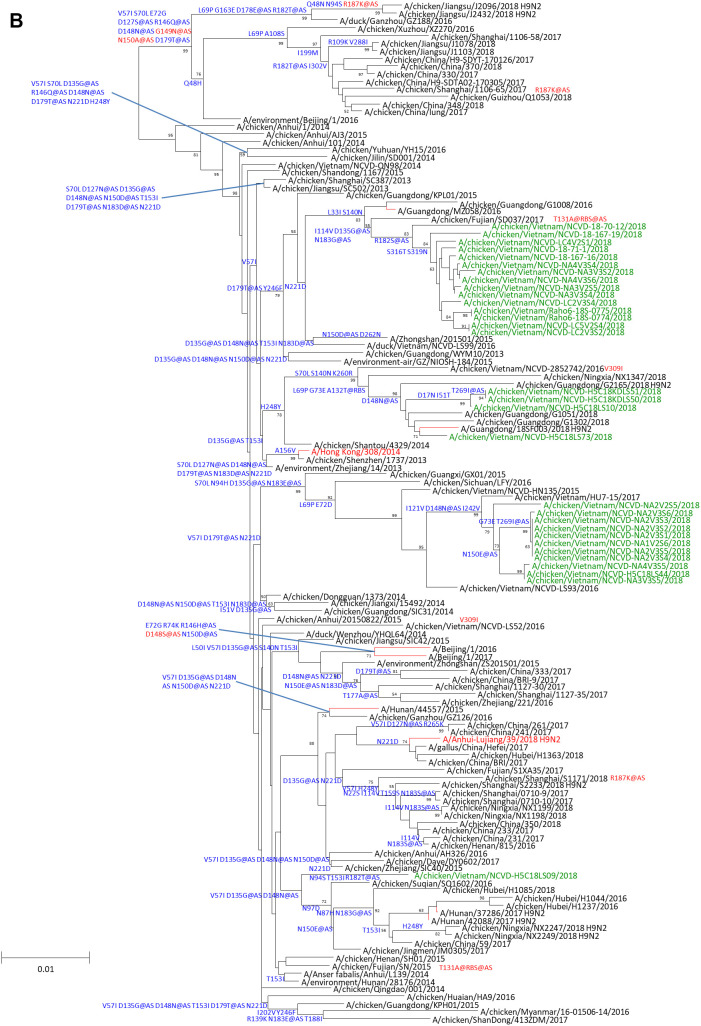
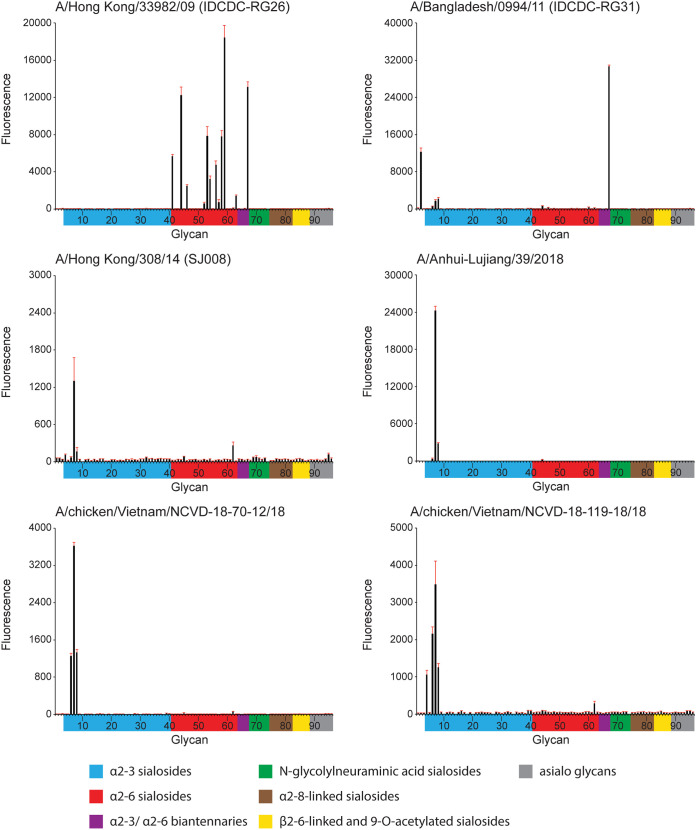
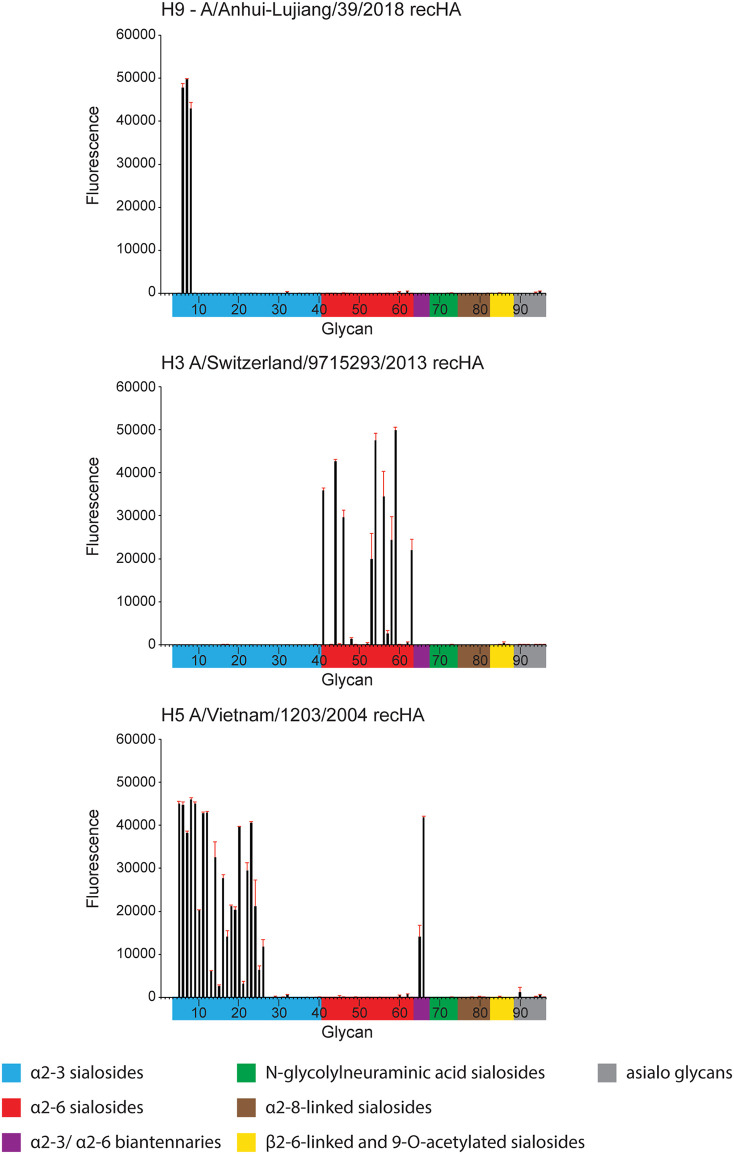
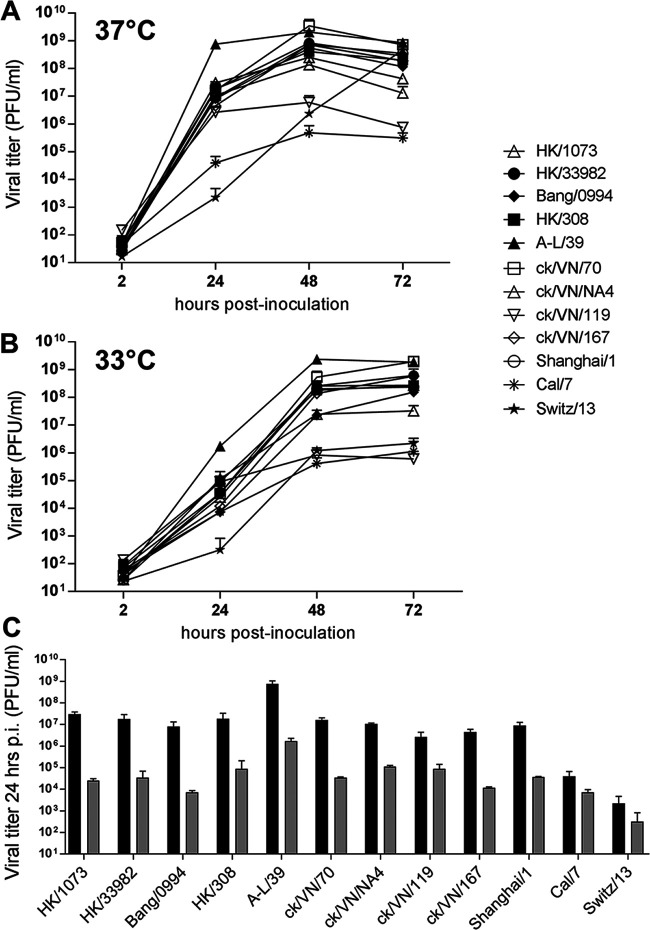
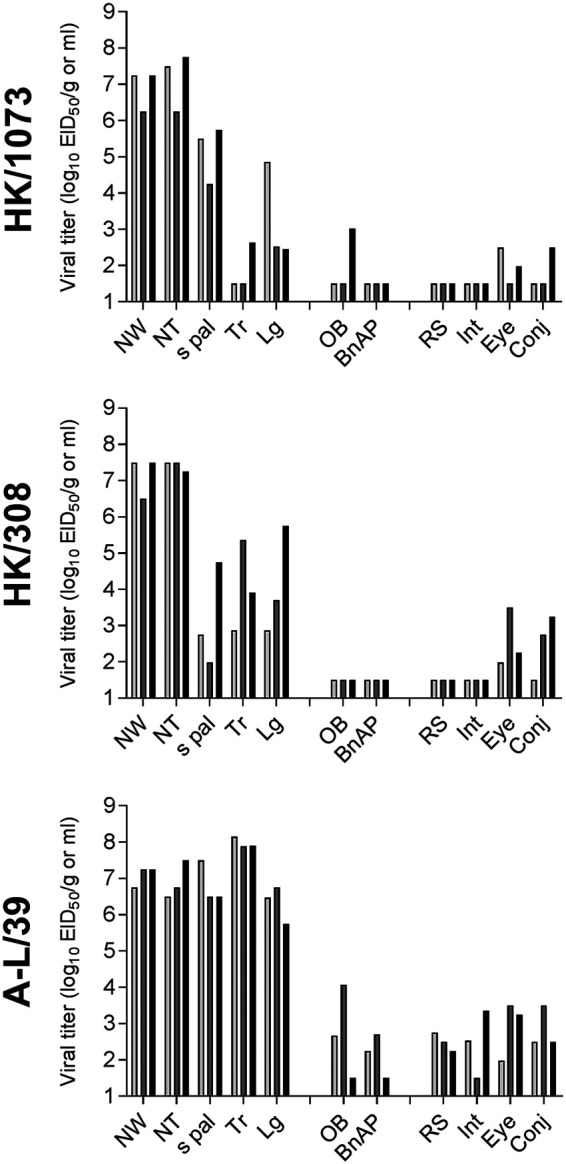
Low-pathogenicity avian influenza A(H9N2) viruses, enzootic in poultry populations in Asia, are associated with fewer confirmed human infections but higher rates of seropositivity compared to A(H5) or A(H7) subtype viruses. Cocirculation of A(H5) and A(H7) viruses leads to the generation of reassortant viruses bearing A(H9N2) internal genes with markers of mammalian adaptation, warranting continued surveillance in both avian and human populations. Here, we describe active surveillance efforts in live poultry markets in Vietnam in 2018 and compare representative viruses to G1 and Y280 lineage viruses that have infected humans. Receptor binding properties, pH thresholds for HA activation, in vitro replication in human respiratory tract cells, and in vivo mammalian pathogenicity and transmissibility were investigated. While A(H9N2) viruses from both poultry and humans exhibited features associated with mammalian adaptation, one human isolate from 2018, A/Anhui-Lujiang/39/2018, exhibited increased capacity for replication and transmission, demonstrating the pandemic potential of A(H9N2) viruses.IMPORTANCE A(H9N2) influenza viruses are widespread in poultry in many parts of the world and for over 20 years have sporadically jumped species barriers to cause human infection. As these viruses continue to diversify genetically and antigenically, it is critical to closely monitor viruses responsible for human infections, to ascertain if A(H9N2) viruses are acquiring properties that make them better suited to infect and spread among humans. In this study, we describe an active poultry surveillance system established in Vietnam to identify the scope of influenza viruses present in live bird markets and the threat they pose to human health. Assessment of a recent A(H9N2) virus isolated from an individual in China in 2018 is also reported, and it was found to exhibit properties of adaptation to humans and, importantly, it shows similarities to strains isolated from the live bird markets of Vietnam.
Keywords: H9N2; influenza; surveillance studies; transmission.
Copyright © 2020 American Society for Microbiology.
Figures
References
-
- Li C, Wang S, Bing G, Carter RA, Wang Z, Wang J, Wang C, Wang L, Wu G, Webster RG, Wang Y, Sun H, Sun Y, Liu J, Pu J. 2017. Genetic evolution of influenza H9N2 viruses isolated from various hosts in China from 1994 to 2013. Emerg Microbes Infect 6:e106. doi: 10.1038/emi.2017.94. - DOI - PMC - PubMed
-
- Li X, Shi J, Guo J, Deng G, Zhang Q, Wang J, He X, Wang K, Chen J, Li Y, Fan J, Kong H, Gu C, Guan Y, Suzuki Y, Kawaoka Y, Liu L, Jiang Y, Tian G, Li Y, Bu Z, Chen H. 2014. Genetics, receptor binding property, and transmissibility in mammals of naturally isolated H9N2 avian influenza viruses. PLoS Pathog 10:e1004508. doi: 10.1371/journal.ppat.1004508. - DOI - PMC - PubMed
-
- Obadan AO, Santos J, Ferreri L, Thompson AJ, Carnaccini S, Geiger G, Gonzalez Reiche AS, Rajao DS, Paulson JC, Perez DR. 2018. Flexibility in vitro of amino acid 226 in the receptor-binding site of an H9 subtype influenza A virus and its effect in vivo on virus replication, tropism, and transmission. J Virol 93:e02011-18. doi: 10.1128/JVI.02011-18. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials
