

Modulation of acoustic navigation behaviour by spatial learning in the echolocating bat Rhinolophus ferrumequinum nippon
- PMID: 32612132
- PMCID: PMC7329871
- DOI: 10.1038/s41598-020-67470-z
Modulation of acoustic navigation behaviour by spatial learning in the echolocating bat Rhinolophus ferrumequinum nippon
Abstract
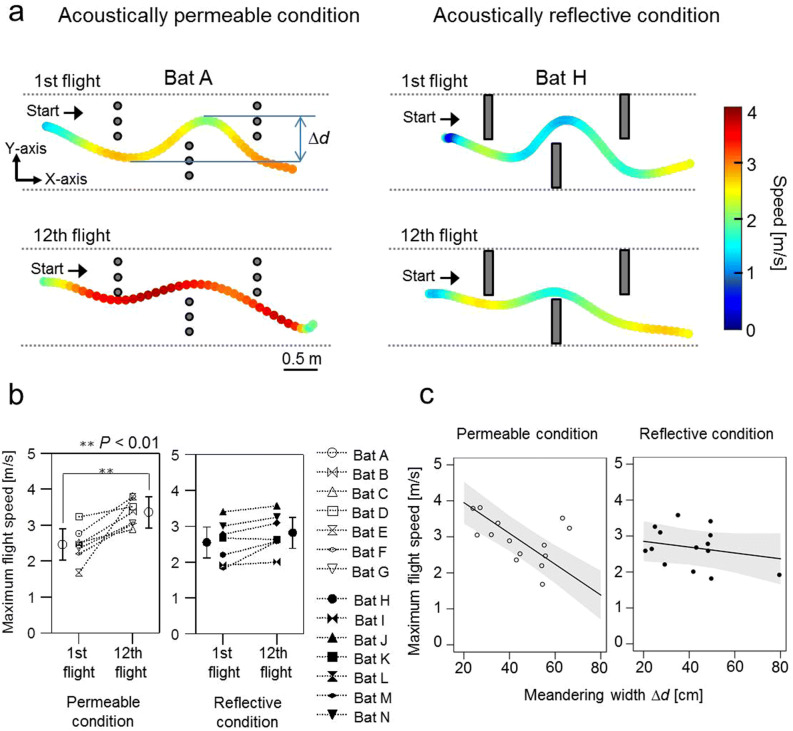
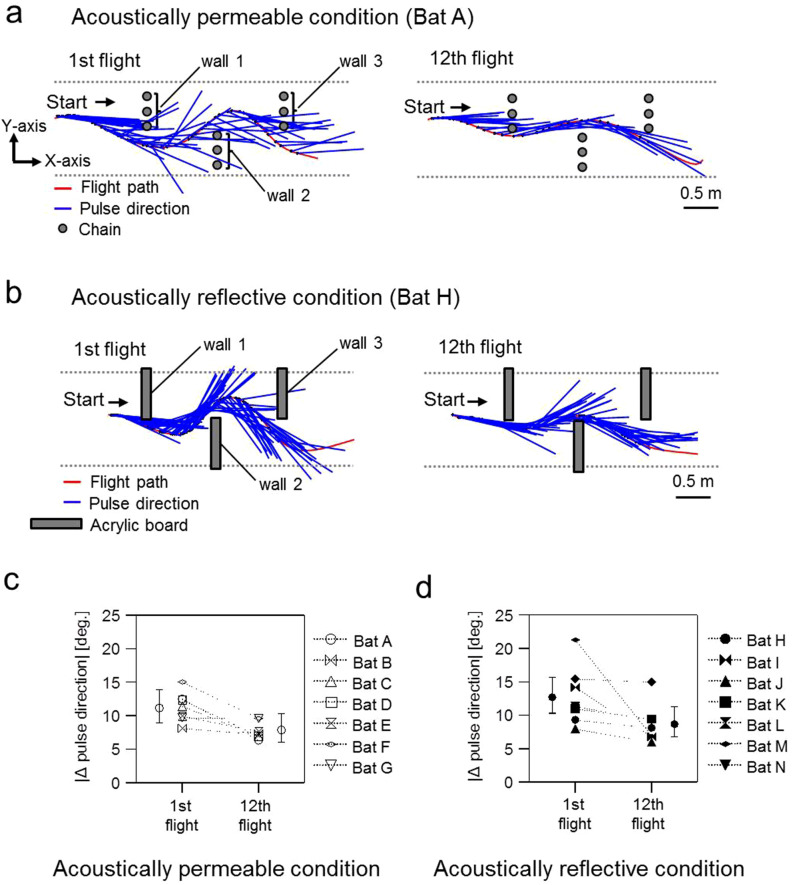
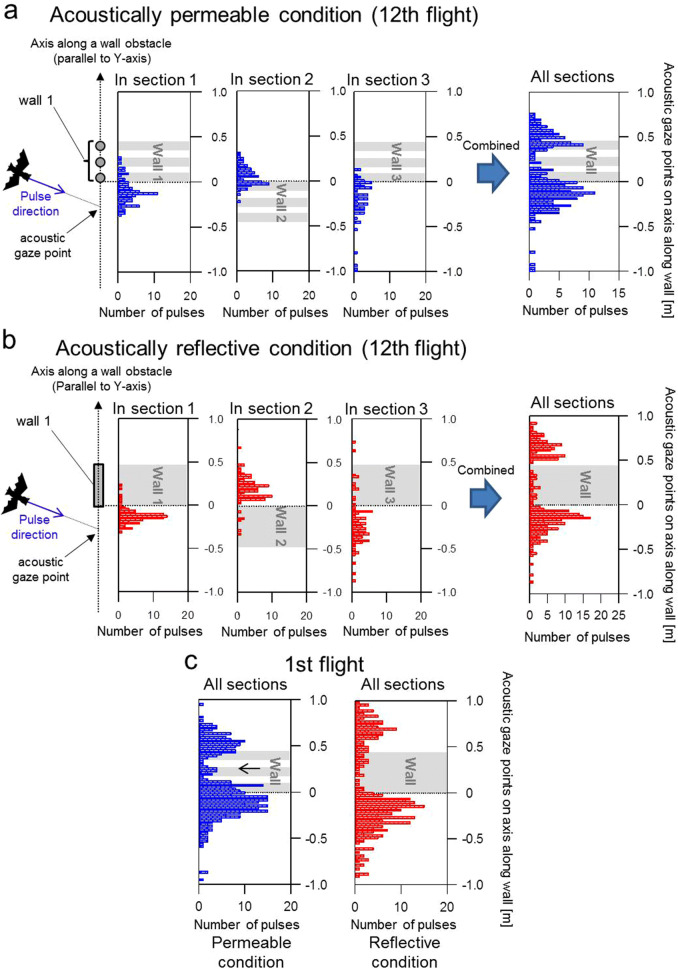
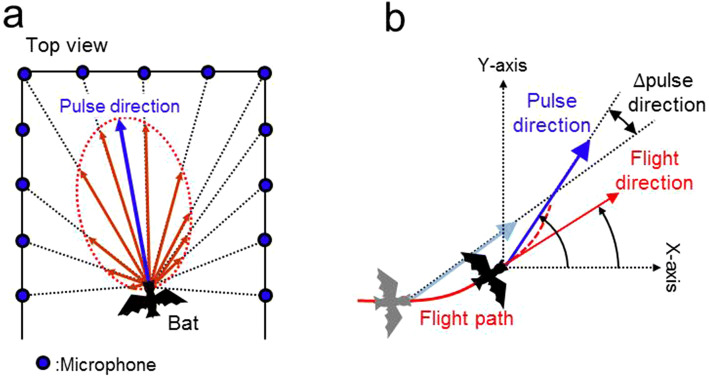
Using echolocation, bats receive acoustic information on their surroundings, which is assumed to help them sophisticatedly navigate complex environments. In this study, to understand spatial learning and acoustic sensing in bats, we investigated how flight and echolocation control changed in Rhinolophus ferrumequinum nippon as they learnt about their surroundings in an obstacle course that they flew through repeatedly. In these experiments, two testing environments (acoustically permeable and acoustically reflective) were prepared using chains and acrylic boards as obstacles to evaluate the interactive effects of spatial learning and flight environments. We found that bats reduced the meandering width of their flights and pulse emissions, and also seemed to reduce their shifts in pulse direction as they learnt more about their environments in both conditions. Throughout all our experiments, the bats with slower flight speeds tended to emit more pulses, which suggests that the number of pulse emissions reflects the echolocation tactics of each bat. The maximum flight speed was especially increased in the acoustically permeable condition, with frequent emissions of multiple pulses (≧triplets) in the early stages of flight, suggesting that bats adjust their flight plan based on how much of their surroundings they are able to sense in advance.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Jen PH-S, Kamada T. Analysis of orientation signals emitted by the CF-FM bat, Pteronotus p. parnellii and the FM bat, Eptesicus fuscus during avoidance of moving and stationary obstacles. J. Comp. Physiol. 1982;148:389–398. doi: 10.1007/BF00679023. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
